Versican



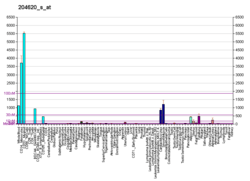

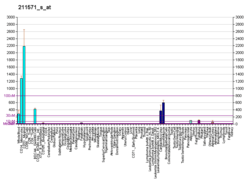
Versican is a large extracellular matrix proteoglycan that is present in a variety of human tissues. It is encoded by the VCAN gene.[5][6]
Versican is a large chondroitin sulfate proteoglycan with an apparent molecular mass of more than 1000kDa. In 1989, Zimmermann and Ruoslahti cloned and sequenced the core protein of fibroblast chondroitin sulfate proteoglycan.[7] They designated it versican in recognition of its versatile modular structure.
Versican belongs to the lectican protein family, with aggrecan (abundant in cartilage), brevican and neurocan (nervous system proteoglycans) as other members. Versican is also known as chondroitin sulfate proteoglycan core protein 2 or chondroitin sulfate proteoglycan 2 (CSPG2), and PG-M.
Structure
These proteoglycans share a homologous globular N-terminal, C-terminal, and glycosaminoglycan (GAG) binding regions.
The N-terminal (G1) globular domain consists of Ig-like loop and two link modules, and has Hyaluronan (HA) binding properties.
Versican occurs in 5 isoforms : V0, V1, V2, V3, V4.[8] The central domain of versican V0 contains both the GAG-α and GAG-β domains. V1 isoforms has the GAG-β domain, V2 has the GAG-α domain, V3 is void of any GAG attachment domains and V4 has a portion of the GAG-β domain. The GAGs, being composed of repeating disaccharide units, contribute to the negative charge and many other properties of proteoglycans.
The C-terminal (G3) globular domain consists of one or two Epidermal growth factor (EGF) repeats, a C-type lectin domain and complement regulatory protein (CRP)-like domain. The C-terminal domain binds a variety of ligands in ECM which contribute significantly to the functions of lecticans.
Function
The role of versican in cell adhesion, migration, and proliferation has been extensively studied. Versican is often considered an anti-adhesion molecule. Considering the large size (>1000 kDa) and hydration capability of versican, it is possible that the interaction of integrins (large family of cell adhesion molecules) with their cell surface receptors is sterically hindered.
Expression of versican is observed in various adult tissues such as blood vessels, skin, and developing heart. Smooth muscle cells of blood vessels, epithelial cells of skin, and the cells of central and peripheral nervous system are a few examples of cell types that express versican physiologically. Versican is involved in development, guiding embryonic cell migration important in the formation of the heart and outlining the path for neural crest cell migration.
N-terminus
The N-terminal of versican has an important role in maintaining the integrity of the ECM by interacting with hyaluronan. Its interactions with link protein has also been studied.
Glycosaminoglycan binding region
The central domain of Versican is decorated with glycosaminoglycans. The structural and functional diversity of Versican is increased by variations in GAG sulfation patterns and the type of GAG chains bound to the core protein. There is a single versican gene, however alternative splicing of its mRNA produces 4 distinct versican isoforms that differ in their potential number of GAG chains. All isoforms have homologous N-terminal (HA binding) and C-terminal (lectin-like) domains. The central domain of versican V0 contains both the GAG-α and GAG-β domains. V1 isoforms has the GAG-β domain, V2 has the GAG-α domain, and V3 is void of any GAG attachment domains, and only consists of the N-terminal and C-terminal globular domains. It is known that the isoforms are differentially expressed in different tissue types. The biological significance of alternative splicing is yet to be determined.
Because of their negatively charged sulfates or carboxyl groups, chondroitin sulfate chains are attracted to various positively charged molecules such as certain growth factors, cytokines, and chemokines. This interaction in the extracellular matrix or on the cell surface is important in the formation of immobilized gradients of these factors, their protection from proteolytic cleavage, and their presentation to specific cell-surface receptors. The binding of versican with leukocyte adhesion molecules L-selectin, P-selectin, and CD44 is also mediated by the interaction of CS chains of versican with the carbohydrate-binding domain of these molecules. Both CD44 and L-selectin have been implicated in leukocyte trafficking. The ability of versican to bind a large panel of chemokines and the biological consequences of such binding has also been examined. Versican can bind specific chemokines through its CS chains and this interaction down-regulates the chemokines function. Recently, in light of results that V1 and V2 isoforms of versican have opposite effects on cell proliferation, glycosaminoglycan domain GAG-β has been implicated in versican-enhanced cell proliferation and versican-induced reduction of cell apoptosis.
C-terminus
The C-terminal of Versican interacts with a variety of molecules in the matrix. One important family of ligands is the tenascin family.[9] For example, The C-lectin domain of versican interacts with tenascin R through its fibronectin type III (FnIII) repeat 3-5 domain in a calcium dependent manner, in vivo. Different tenascin domains interact with a wide range of cellular receptors, including integrins, cell adhesion molecules and members of the syndecan and glypican proteoglycan families. Versican’s C-terminal domain interacts with fibulin-2, a protein whose expression is associated with that of versican in the developing heart. The EGF domain of the C-terminal of versican also binds the EGF-receptor molecule in vivo.
Clinical significance
Versican is a key factor in inflammation through interactions with adhesion molecules on the surfaces of inflammatory leukocytes and interactions with chemokines that are involved in recruiting inflammatory cells.
In the adult central nervous system, versican is found in perineuronal nets, where it may stabilize synaptic connections. Versican can also inhibit nervous system regeneration and axonal growth following an injury to the central nervous system.
Cancer and metastasis
Increased versican expression is often observed in tumor growth in tissues such as breast, brain,[10] ovary, gastrointestinal tract, prostate, and melanoma, sarcoma, and peritoneal mesothelioma. A fifth isoform of versican, V4, that is similar to V1 but with a shortened beta-GAG region, is present and upregulated in human breast cancer.[8]
Versican is required for Lewis lung carcinoma in mice to metastasize to lung, liver and adrenal glands, acting via TLR2 to activate myeloid cells and produce TNF-alpha.[11]
Lung disorders
Versican is increased in the changing tissue extracellular matrix in inflammatory lung disorders such as chronic obstructive pulmonary disease (COPD), asthma and bronchiolitis obliterans syndrome (BOS).[12] Cells (myofibroblasts, macrophages and other inflammatory cells) can migrate more easily through extracellular matrix that has a higher versican content.
Skin disorders
Deposits of versican are not present in normal skin but are found in the reticular dermis during keloid scarring, a condition where scar formation becomes uncontrolled and overgrowth of skin tissue occurs at the site of the wound.[13]
Interactions
Versican has been shown to interact with hyaluronan and a link protein (hyaluronan and proteoglycan link protein 1; HAPLN1).[14]
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000038427 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000021614 - Ensembl, May 2017
- ↑ "Human PubMed Reference:".
- ↑ "Mouse PubMed Reference:".
- ↑ "Entrez Gene: VCAN versican".
- ↑ Iozzo RV, Naso MF, Cannizzaro LA, Wasmuth JJ, McPherson JD (1992). "Mapping of the versican proteoglycan gene (CSPG2) to the long arm of human chromosome 5 (5q12-5q14)". Genomics. 14 (4): 845–51. PMID 1478664. doi:10.1016/S0888-7543(05)80103-X.
- ↑ Zimmermann DR, Ruoslahti E (October 1989). "Multiple domains of the large fibroblast proteoglycan, versican". EMBO J. 8 (10): 2975–81. PMC 401368
 . PMID 2583089.
. PMID 2583089. - 1 2 Kischel P, Waltregny D, Dumont B, Turtoi A, Greffe Y, Kirsch S, De Pauw E, Castronovo V (Feb 2010). "Versican overexpression in human breast cancer lesions: known and new isoforms for stromal tumor targeting". International Journal of Cancer. Journal International Du Cancer. 126 (3): 640–50. PMID 19662655. doi:10.1002/ijc.24812.
- ↑ Aspberg A, Miura R, Bourdoulous S, Shimonaka M, Heinegârd D, Schachner M, Ruoslahti E, Yamaguchi Y (1997). "The C-type lectin domains of lecticans, a family of aggregating chondroitin sulfate proteoglycans, bind tenascin-R by protein-protein interactions independent of carbohydrate moiety". Proc. Natl. Acad. Sci. U.S.A. 94 (19): 10116–21. PMC 23322 . PMID 9294172. doi:10.1073/pnas.94.19.10116.
- ↑ Paulus W, Baur I, Dours-Zimmermann MT, Zimmermann DR (1996). "Differential expression of versican isoforms in brain tumors". J. Neuropathol. Exp. Neurol. 55 (5): 528–33. PMID 8627343. doi:10.1097/00005072-199605000-00005.
- ↑ Kim S, Takahashi H, Lin WW, Descargues P, Grivennikov S, Kim Y, Luo JL, Karin M (Jan 2009). "Carcinoma-produced factors activate myeloid cells through TLR2 to stimulate metastasis". Nature. 457: 102–6. PMC 2746432 . PMID 19122641. doi:10.1038/nature07623.
- ↑ Andersson-Sjöland A, Hallgren O, Rolandsson S, Weitoft M, Tykesson E, Larsson-Callerfelt AK, Rydell-Törmänen K, Bjermer L, Malmström A, Karlsson JC, Westergren-Thorsson G (2015). "Versican in inflammation and tissue remodeling: the impact on lung disorders". Glycobiol. 25 (3): 243–251. PMC 4310351 . PMID 25371494. doi:10.1093/glycob/cwu120.
- ↑ Jumper N, Paus R, Bayat A (22 Apr 2015). "Functional histopathology of keloid disease". Histol. Histopathol. 30 (11624): 1033–57. PMID 25900252. doi:10.14670/HH-11-624.
- ↑ Matsumoto K, Shionyu M, Go M, Shimizu K, Shinomura T, Kimata K, Watanabe H (Oct 2003). "Distinct interaction of versican/PG-M with hyaluronan and link protein". J. Biol. Chem. 278 (42): 41205–12. PMID 12888576. doi:10.1074/jbc.M305060200.
Further reading
- Wight TN, Merrilees MJ (2004). "Proteoglycans in atherosclerosis and restenosis: key roles for versican.". Circ. Res. 94 (9): 1158–67. PMID 15142969. doi:10.1161/01.RES.0000126921.29919.51.
- Kenagy RD, Plaas AH, Wight TN (2006). "Versican degradation and vascular disease.". Trends Cardiovasc. Med. 16 (6): 209–15. PMC 3169384 . PMID 16839865. doi:10.1016/j.tcm.2006.03.011.
- Perides G, Rahemtulla F, Lane WS, Asher RA, Bignami A (1992). "Isolation of a large aggregating proteoglycan from human brain.". J. Biol. Chem. 267 (33): 23883–7. PMID 1429726.
- Iozzo RV, Naso MF, Cannizzaro LA, Wasmuth JJ, McPherson JD (1993). "Mapping of the versican proteoglycan gene (CSPG2) to the long arm of human chromosome 5 (5q12-5q14).". Genomics. 14 (4): 845–51. PMID 1478664. doi:10.1016/S0888-7543(05)80103-X.
- Perides G, Lane WS, Andrews D, Dahl D, Bignami A (1989). "Isolation and partial characterization of a glial hyaluronate-binding protein.". J. Biol. Chem. 264 (10): 5981–7. PMID 2466833.
- Bignami A, Lane WS, Andrews D, Dahl D (1989). "Structural similarity of hyaluronate binding proteins in brain and cartilage.". Brain Res. Bull. 22 (1): 67–70. PMID 2469524. doi:10.1016/0361-9230(89)90129-9.
- Zimmermann DR, Ruoslahti E (1990). "Multiple domains of the large fibroblast proteoglycan, versican.". EMBO J. 8 (10): 2975–81. PMC 401368 . PMID 2583089.
- Krusius T, Gehlsen KR, Ruoslahti E (1987). "A fibroblast chondroitin sulfate proteoglycan core protein contains lectin-like and growth factor-like sequences.". J. Biol. Chem. 262 (27): 13120–5. PMID 2820964.
- Naso MF, Zimmermann DR, Iozzo RV (1995). "Characterization of the complete genomic structure of the human versican gene and functional analysis of its promoter.". J. Biol. Chem. 269 (52): 32999–3008. PMID 7528742.
- Brown DM, Graemiger RA, Hergersberg M, Schinzel A, Messmer EP, Niemeyer G, Schneeberger SA, Streb LM, Taylor CM, Kimura AE (1995). "Genetic linkage of Wagner disease and erosive vitreoretinopathy to chromosome 5q13-14.". Arch. Ophthalmol. 113 (5): 671–5. PMID 7748141. doi:10.1001/archopht.1995.01100050139045.
- Dours-Zimmermann MT, Zimmermann DR (1995). "A novel glycosaminoglycan attachment domain identified in two alternative splice variants of human versican.". J. Biol. Chem. 269 (52): 32992–8. PMID 7806529.
- Zako M, Shinomura T, Ujita M, Ito K, Kimata K (1995). "Expression of PG-M(V3), an alternatively spliced form of PG-M without a chondroitin sulfate attachment in region in mouse and human tissues.". J. Biol. Chem. 270 (8): 3914–8. PMID 7876137. doi:10.1074/jbc.270.8.3914.
- Yao LY, Moody C, Schönherr E, Wight TN, Sandell LJ (1994). "Identification of the proteoglycan versican in aorta and smooth muscle cells by DNA sequence analysis, in situ hybridization and immunohistochemistry.". Matrix Biol. 14 (3): 213–25. PMID 7921538. doi:10.1016/0945-053X(94)90185-6.
- Bode-Lesniewska B, Dours-Zimmermann MT, Odermatt BF, Briner J, Heitz PU, Zimmermann DR (1996). "Distribution of the large aggregating proteoglycan versican in adult human tissues.". J. Histochem. Cytochem. 44 (4): 303–12. PMID 8601689. doi:10.1177/44.4.8601689.
- Sartipy P, Johansen B, Camejo G, Rosengren B, Bondjers G, Hurt-Camejo E (1996). "Binding of human phospholipase A2 type II to proteoglycans. Differential effect of glycosaminoglycans on enzyme activity.". J. Biol. Chem. 271 (42): 26307–14. PMID 8824283. doi:10.1074/jbc.271.42.26307.
- Sartipy P, Bondjers G, Hurt-Camejo E (1999). "Phospholipase A2 type II binds to extracellular matrix biglycan: modulation of its activity on LDL by colocalization in glycosaminoglycan matrixes.". Arterioscler. Thromb. Vasc. Biol. 18 (12): 1934–41. PMID 9848887. doi:10.1161/01.ATV.18.12.1934.
- Perveen R, Hart-Holden N, Dixon MJ, Wiszniewski W, Fryer AE, Brunner HG, Pinkners AJ, van Beersum SE, Black GC (1999). "Refined genetic and physical localization of the Wagner disease (WGN1) locus and the genes CRTL1 and CSPG2 to a 2- to 2.5-cM region of chromosome 5q14.3.". Genomics. 57 (2): 219–26. PMID 10198161. doi:10.1006/geno.1999.5766.
- Lemire JM, Braun KR, Maurel P, Kaplan ED, Schwartz SM, Wight TN (1999). "Versican/PG-M isoforms in vascular smooth muscle cells.". Arterioscler. Thromb. Vasc. Biol. 19 (7): 1630–9. PMID 10397680. doi:10.1161/01.ATV.19.7.1630.
External links
- GeneReviews/NCBI/NIH/UW entry on VCAN-Related Vitreoretinopathy Includes: Wagner Vitreoretinal Degeneration (Wagner Syndrome), Erosive Vitreoretinopathy (ERVR)
- Versicans at the US National Library of Medicine Medical Subject Headings (MeSH)