Reproductive synchrony
Reproductive synchrony is a term used in evolutionary biology and behavioural ecology. Reproductive synchrony — sometimes termed 'ovulatory synchrony' — may manifest itself as 'breeding seasonality'. Where females undergo regular menstruation, 'menstrual synchrony' is another possible term.
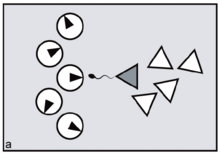
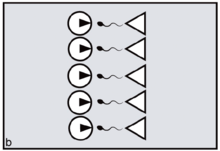
Reproduction is said to be synchronised when fertile matings across a population are temporally clustered, resulting in multiple conceptions (and consequent births) within a restricted time window. In marine and other aquatic contexts, the phenomenon may be referred to as mass spawning. Mass spawning has been observed and recorded in a large number of phyla, including in coral communities within the Great Barrier Reef.[1][2]
In primates, reproductive synchrony usually takes the form of conception and birth seasonality.[3] The regulatory 'clock', in this case, is the sun's position in relation to the tilt of the earth. In nocturnal or partly nocturnal primates — for example, owl monkeys — the periodicity of the moon may also come into play.[4][5] Synchrony in general is for primates an important variable determining the extent of 'paternity skew' — defined as the extent to which fertile matings can be monopolised by a fraction of the population of males. The greater the precision of female reproductive synchrony — the greater the number of ovulating females who must be guarded simultaneously — the harder it is for any dominant male to succeed in monopolising a harem all to himself. This is simply because, by attending to any one fertile female, the male unavoidably leaves the others at liberty to mate with his rivals. The outcome is to distribute paternity more widely across the total male population, reducing paternity skew (figures a, b).[6]
Reproductive synchrony can never be perfect. On the other hand, theoretical models predict that group-living species will tend to synchronise wherever females can benefit by maximising the number of males offered chances of paternity, minimising reproductive skew.[7] For example, the cichlid fish V. moorii spawns in the days leading up to each full moon (lunar synchrony),[8] and broods often exhibit multiple paternity.[9] The same models predict that female primates, including evolving humans, will tend to synchronise wherever fitness benefits can be gained by securing access to multiple males. Conversely, group-living females who need to restrict paternity to a single dominant harem-holder should assist him by avoiding synchrony.[10][11]
In the human case, evolving females with increasingly heavy childcare burdens would have done best by resisting attempts at harem-holding by locally dominant males. No human female needs a partner who will get her pregnant only to disappear, abandoning her in favour of his next sexual partner.[12] To any local group of females, the more such philandering can be successfully resisted — and the greater the proportion of previously excluded males who can be included in the breeding system and persuaded to invest effort — the better.[13] Hence scientists would expect reproductive synchrony — whether seasonal, lunar or a combination of the two — to be central to evolving human strategies of reproductive levelling, reducing paternity skew and culminating in the predominantly monogamous egalitarian norms illustrated by extant hunter-gatherers.[14] Divergent climate regimes differentiating Neanderthal reproductive strategies from those of modern Homo sapiens have recently been analysed in these terms.[15]
See also
References
- ↑ Harrison, P.L.; Babcock, R.C.; Bull, G.D.; Oliver, J.K.; Wallace, C.C.; Willis, B.L. (1984). "Mass Spawning in Tropical Reef Corals". Science. 223 (4641): 1186–1189. PMID 17742935. doi:10.1126/science.223.4641.1186.
- ↑ Babcock, R.C.; Bull, G.D.; Harrison, P.L.; Heyward, A.J.; Oliver, J.K.; Wallace, C.C.; Willis, B.L. (1986). "Synchronous spawnings of 105 scleractinian coral species on the Great Barrier Reef". Marine Biology. 90 (3): 379–394. doi:10.1007/BF00428562.
- ↑ Brockman, D. K. and C. P. Van Schaik, 2005. Seasonality and reproductive function. In D. K. Brockman and C. P. van Schaik (eds), Seasonality in Primates. Studies of living and extinct human and non-human primates. Cambridge: Cambridge University Press, pp. 269-305.
- ↑ Fernandez-Duque, H. de la Iglesa; Erkert, H. G. (2010). "Moonstruck primates: Owl monkeys (Aotus) need moonlight for nocturnal activity in their natural environment". PLoS ONE. 5 (9): e12572. doi:10.1371/journal.pone.0012572.
- ↑ Nash, L. T. 2007. Moonlight and behavior in nocturnal and cathemeral primates, especially Lepilemur leucopus: Illuminating possible anti-predator efforts. In S.L. Gursky and K.A.I. Nekaris (eds), Primate Anti-Predator Strategies. New York: Springer, pp. 173-205.
- ↑ Ostner, J; Nunn, C. L.; Schülke, O. (2008). "Female reproductive synchrony predicts skewed paternity across primates". Behavioral Ecology. 19: 1150–1158. PMC 2583106
 . PMID 19018288. doi:10.1093/beheco/arn093.
. PMID 19018288. doi:10.1093/beheco/arn093. - ↑ Knowlton, N (1979). "Reproductive synchrony, parental investment and the evolutionary dynamics of sexual selection". Animal Behaviour. 27: 1022–33. doi:10.1016/0003-3472(79)90049-6.
- ↑ Rossiter, A (April 1991). "Lunar spawning synchroneity in a freshwater fish". Naturwissenschaften. 78 (4): 182–184. doi:10.1007/bf01136210. Retrieved 1 October 2013.
- ↑ Sefc, Kristina M.; Karin Mattersdorfer; Christian Sturmbauer; Stephan Koblmüller (2008). "High Frequency of Multiple Paternity in Broods of a Socially Monogamous Cichlid Fish with Biparental Nest Defence". Molecular Ecology. 17 (10): 2531–2543. PMID 18430146. doi:10.1111/j.1365-294x.2008.03763.x. Retrieved 30 September 2013.
- ↑ Turke, P. W. (1984). "Effects of ovulatory concealment and synchrony on protohominid mating systems and parental roles". Ethology and Sociobiology. 5: 33–44. doi:10.1016/0162-3095(84)90033-5.
- ↑ Turke, P. W. 1988. Concealed ovulation, menstrual synchrony and paternal investment. In E. Filsinger (ed.), Biosocial Perspectives on the Family. Newbury Park, CA: Sage, pp. 119-136.
- ↑ Power, C. and L. C. Aiello 1997. Female proto-symbolic strategies. In L. D. Hager (ed.), Women in Human Evolution. New York and London: Routledge, pp. 153-171.
- ↑ Bowles, S (2006). "Group competition, reproductive levelling, and the evolution of human altruism". Science. 314: 1569–1572. PMID 17158320. doi:10.1126/science.1134829.
- ↑ Power, C. Power; Arthur, C.; Aiello, L. C. (1997). "On seasonal reproductive synchrony as an evolutionarily stable strategy in human evolution". Current Anthropology. 38 (1): 88–91. doi:10.1086/204586.
- ↑ Power, C.; Sommer, V.; Watts, I. (2013). "The Seasonality Thermostat: Female Reproductive Synchrony and Male Behavior in Monkeys, Neanderthals, and Modern Humans" (PDF). PaleoAnthropology. 2013: 33–60.