Spinosaurus
| Spinosaurus Temporal range: Early–Late Cretaceous, 112–93.5 Ma | |
|---|---|
 | |
| Reconstructed Spinosaurus skeleton in swimming posture | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Order: | Saurischia |
| Suborder: | Theropoda |
| Family: | †Spinosauridae |
| Subfamily: | †Spinosaurinae |
| Genus: | †Spinosaurus Stromer, 1915 |
| Type species | |
| Spinosaurus aegyptiacus Stromer, 1915 | |
| Synonyms | |
| |
Spinosaurus (meaning "spine lizard") is a genus of theropod dinosaur that lived in what now is North Africa, during the upper Albian to upper Turonian stages of the Cretaceous period, about 112 to 93.5 million years ago. This genus was known first from Egyptian remains discovered in 1912 and described by German paleontologist Ernst Stromer in 1915. The original remains were destroyed in World War II, but additional material has come to light in recent years. It is unclear whether one or two species are represented in the fossils reported in the scientific literature. The best known species is S. aegyptiacus from Egypt, although a potential second species, S. maroccanus, has been recovered from Morocco.
Spinosaurus was among the largest of all known carnivorous dinosaurs, nearly as large or even larger than Tyrannosaurus, Giganotosaurus and Carcharodontosaurus. Estimates published in 2005, 2007, and 2008 suggested that it was between 12.6–18 metres (41–59 ft) in length and 7 to 20.9 tonnes (7.7 to 23.0 short tons) in weight.[1][2][3] A new estimate published in 2014 and based on a more complete specimen, supported the earlier research, finding that Spinosaurus could reach lengths of over 15 m (49 ft).[4] The skull of Spinosaurus was long and narrow, similar to that of a modern crocodilian. Spinosaurus is known to have eaten fish, and most scientists believe that it hunted both terrestrial and aquatic prey; evidence suggests that it lived both on land and in water as a modern crocodilian does. The distinctive spines of Spinosaurus, which were long extensions of the vertebrae, grew to at least 1.65 meters (5.4 ft) long and were likely to have had skin connecting them, forming a sail-like structure, although some authors have suggested that the spines were covered in fat and formed a hump. Multiple functions have been put forward for this structure, including thermoregulation and display.
Description
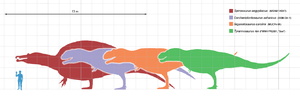
Since its discovery, Spinosaurus has been a contender for the longest and largest theropod dinosaur.[5] Both Friedrich von Huene in 1926[6] and Donald F. Glut in 1982 listed it as among the most massive theropods in their surveys, at 15 meters (49 ft) in length and upwards of 6 t (5.9 long tons; 6.6 short tons) in weight.[7] In 1988, Gregory Paul also listed it as the longest theropod at 15 meters (49 ft), but gave a lower mass estimate of 4 tonnes (3.9 long tons; 4.4 short tons).[8]
Dal Sasso et al. (2005) assumed that Spinosaurus and Suchomimus had the same body proportions in relation to their skull lengths, and thereby calculated that Spinosaurus was 16 to 18 meters (52 to 59 ft) in length and 7 to 9 tonnes (6.9 to 8.9 long tons; 7.7 to 9.9 short tons) in weight.[3] The Dal Sasso et al. estimates were criticized because the skull length estimate was uncertain, and (assuming that body mass increases as the cube of body length) scaling Suchomimus which was 11 meters (36 ft) long and 3.8 tonnes (4.2 short tons) in mass to the range of estimated lengths of Spinosaurus would produce an estimated body mass of 11.7 to 16.7 tonnes (12.9 to 18.4 short tons).[2]

François Therrien and Donald Henderson, in a 2007 paper using scaling based on skull length, challenged previous estimates of the size of Spinosaurus, finding the length too great and the weight too small.[2] Based on estimated skull lengths of 1.5 to 1.75 meters (4.9 to 5.7 ft), their estimates include a body length of 12.6 to 14.3 meters (41 to 47 ft) and a body mass of 12 to 20.9 tonnes (11.8 to 20.6 long tons; 13.2 to 23.0 short tons).[2] The lower estimates for Spinosaurus would imply that the animal was shorter and lighter than Carcharodontosaurus and Giganotosaurus.[2] The Therrien and Henderson study has been criticized for the choice of theropods used for comparison (e.g., most of the theropods used to set the initial equations were tyrannosaurids and carnosaurs, which have a different build than spinosaurids), and for the assumption that the Spinosaurus skull could be as little as 1.5 meters (4.9 ft) in length.[9][10] Improvement of the precision of size estimates for Spinosaurus requires the discovery of more complete remains as available for some other dinosaurs,[11] especially the limb bones of Spinosaurus which are "hitherto unknown".[3]
Neural spines

Very tall neural spines growing on the back vertebrae of Spinosaurus formed the basis of what is usually called the animal's "sail". The lengths of the neural spines reached over 10 times the diameters of the vertebral bodies from which they extended.[12] The neural spines were slightly longer front to back at the base than higher up, and were unlike the thin rods seen in the pelycosaur finbacks Edaphosaurus and Dimetrodon, contrasting also with the thicker spines in the iguanodontian Ouranosaurus.[12]
Spinosaurus sails were unusual, although other dinosaurs, namely the ornithopod Ouranosaurus, which lived a few million years earlier in the same general region as Spinosaurus, and the South American sauropod Amargasaurus, might have developed similar structural adaptations of their vertebrae. The sail may be an analog of the sail of the Permian synapsid Dimetrodon, which lived before the dinosaurs even appeared, produced by convergent evolution.[12]
The structure may also have been more hump-like than sail-like, as noted by Stromer in 1915 ("one might rather think of the existence of a large hump of fat [German: Fettbuckel], to which the [neural spines] gave internal support")[13] and by Jack Bowman Bailey in 1997.[12] In support of his "buffalo-back" hypothesis, Bailey argued that in Spinosaurus, Ouranosaurus, and other dinosaurs with long neural spines, the spines were relatively shorter and thicker than the spines of pelycosaurs (which were known to have sails); instead, the dinosaurs' neural spines were similar to the neural spines of extinct hump-backed mammals such as Megacerops and Bison latifrons.[12]
Skull

The skull had a narrow snout filled with straight conical teeth that lacked serrations. There were six or seven teeth on each side of the very front of the upper jaw, in the premaxillae, and another twelve in both maxillae behind them. The second and third teeth on each side were noticeably larger than the rest of the teeth in the premaxilla, creating a space between them and the large teeth in the anterior maxilla; large teeth in the lower jaw faced this space. The very tip of the snout holding those few large anterior teeth was expanded, and a small crest was present in front of the eyes.[3] Using the dimensions of three specimens known as MSNM V4047, UCPC-2, and BSP 1912 VIII 19, and assuming that the postorbital part of the skull of MSNM V4047 had a shape similar to the postorbital part of the skull of Irritator, Dal Sasso et al. (2005) estimated that the skull of Spinosaurus was 1.75 meters (5.7 ft) long.[3] The Dal Sasso et al. skull length estimate was questioned because skull shapes can vary across spinosaurid species.[2]
A 2013 made study performed by scientists Andrew R. Cuff and Emily Rayfield showed that Spinosaurids like Spinosaurus had relatively poor resistance in their skulls for torsion compared to other members of this group (Baryonyx) and modern alligators, thus showing Spinosaurus preyed more regularly on fish than it did on land animals, although considered predators of the former too.[14]
Discovery and naming
Naming of species

Two species of Spinosaurus have been named: Spinosaurus aegyptiacus (meaning "Egyptian spine lizard") and Spinosaurus maroccanus (meaning "Moroccan spine lizard").[15][16][17]
The first described remains of Spinosaurus were found and described in the early 20th century. In 1912, Richard Markgraf discovered a partial skeleton of a dinosaur in the Bahariya Formation of western Egypt. In 1915, German paleontologist Ernst Stromer published an article assigning the specimen to a new genus and species Spinosaurus aegyptiacus.[13][15]
Fragmentary additional remains from Bahariya, including vertebrae and hindlimb bones, were designated by Stromer as "Spinosaurus B" in 1934.[18] Stromer considered them different enough to belong to another species, and this has been borne out. With the advantage of more expeditions and material, it appears that they pertain either to Carcharodontosaurus[19] or to Sigilmassasaurus.[16]
S. maroccanus was originally described by Dale Russell in 1996 as a new species based on the length of its neck vertebrae.[16] Specifically, Russell claimed that the ratio of the length of the centrum (body of vertebra) to the height of the posterior articular facet was 1.1 in S. aegyptiacus and 1.5 in S. maroccanus.[16] Later authors have been split on this topic. Some authors note that the length of the vertebrae can vary from individual to individual, that the holotype specimen was destroyed and thus cannot be compared directly with the S. maroccanus specimen, and that it is unknown which cervical vertebrae the S. maroccanus specimens represent. Therefore, though some have retained the species as valid without much comment,[20][21][22] most researchers regard S. maroccanus as a nomen dubium[3][23][24] or as a junior synonym of S. aegyptiacus.[19]
Specimens
Six main partial specimens of Spinosaurus have been described.

BSP 1912 VIII 19, described by Stromer in 1915 from the Bahariya Formation, was the holotype.[13][15] The material consisted of the following items, most of which were incomplete: right and left dentaries and splenials from the lower jaw measuring 75 centimeters (30 in) long; a straight piece of the left maxilla that was described but not drawn; 20 teeth; 2 cervical vertebrae; 7 dorsal (trunk) vertebrae; 3 sacral vertebrae; 1 caudal vertebra; 4 thoracic ribs; and gastralia.[13] Of the nine neural spines whose heights are given, the longest ("i," associated with a dorsal vertebra) was 1.65 meters (5.4 ft) in length.[13] Stromer claimed that the specimen was from the early Cenomanian, approximately 97 million years ago.[13][15]
This specimen was destroyed in World War II, specifically "during the night of 24/25 April 1944 in a British bombing raid of Munich" that severely damaged the building housing the Paläontologisches Museum München (Bavarian State Collection of Paleontology).[15] However, detailed drawings and descriptions of the specimen remain. Stromer's son donated Stromer's archives to the Paläontologische Staatssammlung München in 1995, and Smith et al. analyzed two photographs of the Spinosaurus holotype specimen BSP 1912 VIII 19 discovered in the archives in 2000.[15] On the basis of a photograph of the lower jaw and a photograph of the entire specimen as mounted, Smith concluded that Stromer's original 1915 drawings were slightly inaccurate.[15] In 2003, Oliver Rauhut suggested that Stromer's Spinosaurus holotype was a chimera, composed of vertebrae and neural spines from a carcharodontosaurid similar to Acrocanthosaurus and a dentary from Baryonyx or Suchomimus.[24] This analysis was rejected in at least one subsequent paper.[3]
NMC 50791, held by the Canadian Museum of Nature, is a mid-cervical vertebra which is 19.5 centimeters (7.7 in) long from the Kem Kem Beds of Morocco.[16] It is the holotype of Spinosaurus maroccanus as described by Russell in 1996.[16] Other specimens referred to S. maroccanus in the same paper were two other mid-cervical vertebrae (NMC 41768 and NMC 50790), an anterior dentary fragment (NMC 50832), a mid-dentary fragment (NMC 50833), and an anterior dorsal neural arch (NMC 50813).[16] Russell stated that "only general locality information could be provided" for the specimen, and therefore it could be dated only "possibly" to the Albian.[16]
MNHN SAM 124, housed at the Muséum National d'Histoire Naturelle, is a snout (consisting of partial premaxillae, partial maxillae, vomers, and a dentary fragment).[25] Described by Taquet and Russell in 1998, the specimen is 13.4 to 13.6 centimeters (5.3–5.4 in) in width; no length was stated.[25] The specimen was located in Algeria, and "is of Albian age."[25] Taquet and Russell believed that this specimen along with a premaxilla fragment (SAM 125), two cervical vertebrae (SAM 126-127), and a dorsal neural arch (SAM 128), belonged to S. maroccanus.[25]
BM231 (in the collection of the Office National des Mines, Tunis) was described by Buffetaut and Ouaja in 2002.[23] It consists of a partial anterior dentary 11.5 centimetres (4.53 in) in length from an early Albian stratum of the Chenini Formation of Tunisia.[23] The dentary fragment, which included four alveoli and two partial teeth, was "extremely similar" to existing material of S. aegyptiacus.[23]
UCPC-2 in the University of Chicago Paleontological Collection consists mainly of two narrow connected nasals with a "fluted crest" from the region between the eyes.[3] The specimen, which is 18.0 centimetres (7.09 in) long, was located in an early Cenomanian part of the Moroccan Kem Kem Beds in 1996 and described in the scientific literature in 2005 by Cristiano Dal Sasso of the Civic Natural History Museum in Milan and colleagues.[3]

MSNM V4047 (in the Museo di Storia Naturale di Milano), described by Dal Sasso et al. in 2005, consists of a snout (premaxillae, partial maxillae, and partial nasals) 98.8 centimetres (38.9 in) long from the Kem Kem Beds.[3] Like UCPC-2, it is thought to have come from the early Cenomanian.
FSAC-KK 11888 is a partial subadult skeleton recovered from the Kem Kem beds of North Africa. Described by Ibrahim et al. (2014) and designated as the neotype specimen (although Evers et al. 2015 reject the neotype designation for FSAC-KK-11888).[26] It includes cervical vertebrae, dorsal vertebrae, neural spines, a complete sacrum, femora, tibiae, pedal phalanges, caudal vertebra, several dorsal ribs, and fragments of the skull.[27] The body proportions of this specimen have been debated as the hind limbs are disproportionately shorter in the specimen than in previous reconstructions. However, it has been demonstrated by multiple paleontologists that the specimen is not a chimaera, and is indeed a specimen of Spinosaurus that suggests that the animal had much smaller hind limbs than previously thought [28][29][30] Other known specimens consist mainly of very fragmentary remains and scattered teeth. These include:
- A 1986 paper described prismatic structures in tooth enamel from two Spinosaurus teeth from Tunisia.[31]
- Buffetaut (1989, 1992) referred three specimens from the Institut und Museum für Geologie und Paläontologie of the University of Göttingen in Germany to Spinosaurus: a right maxilla fragment IMGP 969-1, a jaw fragment IMGP 969-2, and a tooth IMGP 969-3.[32][33] These had been found in a Lower Cenomanian or Upper Albian deposit in southeastern Morocco in 1971.[32]

- Kellner and Mader (1997) described two unserrated spinosaurid teeth from Morocco (LINHM 001 and 002) that were "highly similar" to the teeth of the S. aegyptiacus holotype.[34]
- Teeth from the Chenini Formation in Tunisia which are "narrow, somewhat rounded in cross-section, and lack the anterior and posterior serrated edges characteristic of theropods and basal archosaurs" were assigned to Spinosaurus in 2000.[35]
- Teeth from the Echkar Formation of Niger were "tentatively" referred to Spinosaurus in 2007.[36]
- A partial tooth 8 cm long purchased at a fossil trade show, reportedly from the Kem Kem Bed of Morocco and attributed to Spinosaurus maroccanus, showed 1–5 mm wide longitudinal striations and micro-structures (irregular ridges) among the striations in a 2010 paper.[22]
MHNM.KK374 to.KK378 are five isolated quadrates (skull bones) of different sizes were collected by locals and acquired commercially in the Kem Kem region of southeastern Morocco, provided by François Escuillié and are deposited in the collections of the Muséum d’Histoire Naturelle of Marrakech. The quadrates show two different morphologies suggesting the existence of two spinosaurines in Morocco.[37]
Possible specimens
Possible material belonging to Spinosaurus has been reported from the Turkana Grits of Kenya.[38]
Some scientists have considered the genus Sigilmassasaurus a junior synonym of Spinosaurus. In Ibrahim et al. (2014), the specimens of Sigilmassasaurus was referred to Spinosaurus aegyptiacus together with "Spinosaurus B" as the neotype and Spinosaurus maroccanus was considered as a nomen dubium following the conclusions of the other papers.[3][4][19] A 2015 re-description of Sigilmassasaurus disputed these conclusions, and considered the genus valid.[26]
Classification


Spinosaurus gives its name to the Spinosauridae family of dinosaurs, which includes two subfamilies: Baryonychinae and Spinosaurinae. The Baryonychinae include Baryonyx from southern England and Suchomimus from Niger in central Africa. The Spinosaurinae include Spinosaurus, Irritator from Brazil, and Angaturama (which is probably synonymous with Irritator) from Brazil.[3] The Spinosaurinae share unserrated straight teeth that are widely spaced (e.g., 12 on one side of the maxilla), as opposed to the Baryonychinae which have serrated curved teeth that are numerous (e.g., 30 on one side of the maxilla).[3][20]
The following cladogram shows an analysis of Tetanurae simplified to show only Spinosauridae from Allain et al. (2012):[39]
| Spinosauridae |
| ||||||||||||||||||||||||
Paleobiology
Function of neural spines

The function of the dinosaur's sail or hump is uncertain; scientists have proposed several hypotheses including heat regulation and display. In addition, such a prominent feature on its back could make it appear even larger than it was, intimidating other animals.[12]
The structure may have been used for thermoregulation. If the structure contained abundant blood vessels, the animal could have used the sail's large surface area to absorb heat. This would imply that the animal was only partly warm-blooded at best and lived in climates where nighttime temperatures were cool or low and the sky usually not cloudy. It is also possible that the structure was used to radiate excess heat from the body, rather than to collect it. Large animals, due to the relatively small ratio of surface area of their body compared to the overall volume (Haldane's principle), face far greater problems of dissipating excess heat at higher temperatures than gaining it at lower. Sails of large dinosaurs added considerably to the skin area of their bodies, with minimum increase of volume. Furthermore, if the sail was turned away from the sun, or positioned at a 90 degree angle towards a cooling wind, the animal would quite effectively cool itself in the warm climate of Cretaceous Africa.[40] However, Bailey (1997) was of the opinion that a sail could have absorbed more heat than it radiated.[12] Bailey proposed instead that Spinosaurus and other dinosaurs with long neural spines had fatty humps on their backs for energy storage, insulation, and shielding from heat.[12]
Elaborate body structures of many modern-day animals usually serve to attract members of the opposite sex during mating. It is quite possible that the sails or humps of these dinosaurs were used for courtship, in a way similar to a peacock's tail. Stromer speculated that males and females may have differed in the size of the neural spine.[13]
Gimsa et al. (2015) suggest that the dorsal sail of Spinosaurus was homologous to the dorsal fins of sailfish and served a hydrodynamic purpose.[41] Gimsa and others point out that more basal, long-legged spinosaurids have otherwise round or crescent-shaped dorsal sails, whereas in Spinosaurus, the dorsal neural spines form a shape that is roughly rectangular and similar in shape to the dorsal fins of sailfish. They therefore argue that Spinosaurus used its dorsal neural sail in the same manner as Sailfish, and that it also employed its long narrow tail to stun prey like a modern Thresher shark. Sailfish employ their dorsal fins for herding schools of fish into a "bait hall" where they cooperate to trap the fish into a certain area where the sailfish can snatch the fish with their bills. The sail could have possibly reduced yaw rotation by counteracting the lateral force in the direction opposite to the slash as suggested by Gimsa et al. (2015). Gimsa and colleagues specifically wrote :

Spinosaurus anatomy exhibits another feature that may have a modern homology: its long tail resembled that of the thresher shark, employed to slap the water to herd and stun shoals of fish before devouring them (Oliver et al.2013). The strategies that sailfish and thresher sharks employ against shoaling fish are more effective when the shoal is first concentrated into a ‘bait ball’ (Helfman, Collette & Facey, 1997; Oliver et al.2013; Domenici et al.2014). Since this is difficult for individual predators to achieve, they cooperate in this effort. When herding a shoal of fish or squid, sailfish also raise their sails to make themselves appear larger. When they slash or wipe their bills through shoaling fish by turning their heads, their dorsal sail and fins are outstretched to stabilize their bodies hydrodynamically (Lauder & Drucker, 2004). Domenici et al. (2014) postulate that these fin extensions enhance the accuracy of tapping and slashing. The sail can reduce yaw rotation by counteracting the lateral force in the direction opposite to the slash. This means that prey is less likely to recognize the massive trunk as being part of an approaching predator (Marras et al.2015; Webb & Weihs 2015). Film footage available online impressively demonstrates the hunting strategies of sailfish and thresher sharks. Interestingly, Spinosaurus exhibited the anatomical features required to combine all three hunting strategies: a sail for herding prey more efficiently, as well as flexible tail and neck to slap the water for stunning, injuring or killing prey. The submerged dorsal sail would have provided a strong centreboard-like counterforce for powerful sidewards movements of the strong neck and long tail, as performed by sailfish (Domenici et al.2014) or thresher sharks (Oliver et al.2013). While smaller dorsal sails or fins make the dorsal water volume better accessible for slashing, it can be speculated that their smaller stabilization effect makes lateral slashing less efficient (e.g. for thresher sharks). Forming a hydrodynamic fulcrum and hydrodynamically stabilizing the trunk along the dorsoventral axis, Spinosaurus’ sail would also have compensated for the inertia of the lateral neck by tail movements and vice versa not only for predation but also for accelerated swimming. This behaviour might also have been one reason for Spinosaurus’ muscular chest and neck reported by Ibrahim et al. (2014). " Finally, it is quite possible that the sail or hump combined these functions, acting normally as a heat regulator, becoming a courting aid during the mating season, being used to cool itself and, on occasions, turning into an intimidating device when an animal was feeling threatened.[12]
Diet

It is unclear whether Spinosaurus was primarily a terrestrial predator or a piscivore, as indicated by its elongated jaws, conical teeth and raised nostrils. The hypothesis of spinosaurs as specialized fish eaters has been suggested before by A. J. Charig and A. C. Milner for Baryonyx. They base this on the anatomical similarity with crocodilians and the presence of digestive acid-etched fish scales in the rib cage of the type specimen.[42] Large fish are known from the faunas containing other spinosaurids, including the Mawsonia, in the mid-Cretaceous of northern Africa and Brazil. Direct evidence for spinosaur diet comes from related European and South American taxa. Baryonyx was found with fish scales and bones from juvenile Iguanodon in its stomach, while a tooth embedded in a South American pterosaur bone suggests that spinosaurs occasionally preyed on pterosaurs,[43] but Spinosaurus was likely to have been a generalized and opportunistic predator, possibly a Cretaceous equivalent of large grizzly bears, being biased toward fishing, though it undoubtedly scavenged and took many kinds of small or medium-sized prey.[8] A study by Cuff and Rayfield (2013) concluded that bio-mechanical data suggests that Spinosaurus was not an obligate piscivore and that its diet was more closely associated with each individual's size.[14] The characteristic rostral morphology of Spinosaurus allowed its jaws to resist bending in the vertical direction, however its jaws were poorly adapted with respect to resisting lateral bending.[14]
In 2009, Dal Sasso et al.. reported the results of X-ray computed tomography of the MSNM V4047 snout.[44] As the foramina on the outside all communicated with a space on the inside of the snout, the authors speculated that Spinosaurus had pressure receptors inside the space that allowed it to hold its snout at the surface of the water to detect swimming prey species without seeing them.[44]
A 2010 isotope analysis by Romain Amiot and colleagues found that oxygen isotope ratios of spinosaurid teeth, including teeth of Spinosaurus, indicate semiaquatic lifestyles.[45] Isotope ratios from tooth enamel and from other parts of Spinosaurus (found in Morocco and Tunisia) and of other predators from the same area such as Carcharodontosaurus were compared with isotopic compositions from contemporaneous theropods, turtles, and crocodilians.[45] The study found that Spinosaurus teeth from five of six sampled localities had oxygen isotope ratios closer to those of turtles and crocodilians when compared with other theropod teeth from the same localities.[45] The authors postulated that Spinosaurus switched between terrestrial and aquatic habitats to compete for food with large crocodilians and other large theropods respectively.[45]
Posture


Although traditionally depicted as a biped, it has been suggested since the mid-1970s that Spinosaurus was at least an occasional quadruped.[7][40] This has been bolstered by the discovery of Baryonyx, a relative with robust arms.[46] Because of the mass of the hypothesized fatty dorsal humps of Spinosaurus, Bailey (1997) was open to the possibility of a quadrupedal posture,[12] leading to new restorations of it as such.[46] The hypothesis that Spinosaurus had a typical quadrupedal gait has fallen out of favor, though spinosaurids may have crouched in a quadrupedal posture.[42]
Theropods, including spinosaurids, could not pronate their hands (rotate the forearm so the palm faced the ground),[47] but a resting position on the side of the hand was possible, as shown by fossil prints from an Early Jurassic theropod.[48] A 2014 paper describing new material of Spinosaurus, proposed that its legs were too short for it to move effectively on land. The reconstruction used in the study was an extrapolation based on different sized individuals, scaled to what was assumed to be the correct proportions.[49] Palaeontologist John Hutchinson of the Royal Veterinary College of the University of London has expressed scepticism to the new reconstruction, and cautioned that using different specimens can result in inaccurate chimaeras.[50] Scott Hartman also expressed criticism because he believes the legs and the pelvis were inaccurately scaled (27% too short) and don't match the published lengths.[51] However, responses from Ibrahim et al. to Mark Witton have been positively received as reliable.[52] The 2015 re-description of Sigilmassasaurus[26] Evers et al. 2015 doubted whether the material assigned to Spinosaurus by Ibrahim et. al. belonged to it.[26]
Paleoecology
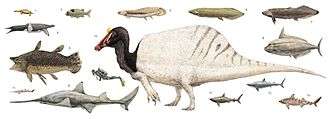
The environment inhabited by Spinosaurus is only partially understood, and covers a great deal of what is now northern Africa. The region of Africa Spinosaurus is preserved in dates from 112 to 93.5 million years ago.[53][15][1] A 1996 study concluded from Moroccan fossils that Spinosaurus, Carcharodontosaurus, and Deltadromeus "ranged across north Africa during the late Cretaceous (Cenomanian)."[54] Those Spinosaurus that lived in the Bahariya Formation of what is now Egypt may have contended with shoreline conditions on tidal flats and channels, living in mangrove forests alongside similarly large dinosaurian predators Bahariasaurus and Carcharodontosaurus, the titanosaur sauropods Paralititan and Aegyptosaurus, crocodylomorphs, bony and cartilaginous fish, turtles, lizards, and plesiosaurs.[55] In the dry season it might have resorted to preying on pterosaurs.[56] This situation resembles that in the Late Jurassic Morrison Formation of North America, which boasts up to five theropod genera over one tonne in weight, as well as several smaller genera (Henderson, 1998; Holtz et al., 2004). Differences in head shape and body size among the large North African theropods may have been enough to allow niche partitioning as seen among the many different predator species found today in the African savanna (Farlow & Pianka, 2002).
In popular culture

Spinosaurus appeared in the 2001 film Jurassic Park III, replacing Tyrannosaurus as the main antagonist.[57] The film's consulting paleontologist John R. Horner was quoted as saying: "If we base the ferocious factor on the length of the animal, there was nothing that ever lived on this planet that could match this creature [Spinosaurus]. Also my hypothesis is that T-rex was actually a scavenger rather than a killer. Spinosaurus was really the predatory animal."[58] (He has since retracted the statement about T. Rex being a scavenger.) In the film, Spinosaurus was portrayed as larger and more powerful than Tyrannosaurus: in a scene depicting a battle between the two resurrected predators, Spinosaurus emerges victorious by snapping the tyrannosaur's neck.[59] In the fourth film Jurassic World, there is a nod to this fight where the T-Rex smashes through the skeleton of a Spinosaurus in the climatic fight near the end of the film.[60]
Spinosaurus has long been depicted in popular books about dinosaurs, although only recently has there been enough information about spinosaurids for an accurate depiction. After an influential 1955 skeletal reconstruction by Lapparent and Lavocat[61] based on a 1936 diagram by Stromer,[62] it has been treated as a generalized upright theropod, with a skull similar to that of other large theropods and a sail on its back, even having four-fingered hands.[46]
In addition to films, action figures, video games, and books, Spinosaurus has been depicted on postage stamps such as ones from Angola, The Gambia, and Tanzania.[63][64]
References
- 1 2 Holtz, Thomas R. Jr. (2012) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages, Winter 2011 Appendix.
- 1 2 3 4 5 6 Therrien, F.; Henderson, D.M. (2007). "My theropod is bigger than yours...or not: estimating body size from skull length in theropods". Journal of Vertebrate Paleontology. 27 (1): 108–115. ISSN 0272-4634. doi:10.1671/0272-4634(2007)27[108:MTIBTY]2.0.CO;2.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 dal Sasso, C.; Maganuco, S.; Buffetaut, E.; Mendez, M.A. (2005). "New information on the skull of the enigmatic theropod Spinosaurus, with remarks on its sizes and affinities". Journal of Vertebrate Paleontology. 25 (4): 888–896. ISSN 0272-4634. doi:10.1671/0272-4634(2005)025[0888:NIOTSO]2.0.CO;2.
- 1 2 Ibrahim, Nizar; Sereno, Paul C.; Dal Sasso, Cristiano; Maganuco, Simone; Fabri, Matteo; Martill, David M.; Zouhri, Samir; Myhrvold, Nathan; Lurino, Dawid A. (2014). "Semiaquatic adaptations in a giant predatory dinosaur". Science. 345 (6204): 1613–6. PMID 25213375. doi:10.1126/science.1258750. Supplementary Information
- ↑ nationalgeographic.com 'River Monster': 50-Foot Spinosaurus
- ↑ von Huene, F.R. (1926). "The carnivorous saurischia in the Jura and Cretaceous formations principally in Europe". Rev. Mus. La Plata. 29: 35–167.
- 1 2 Glut, D.F. (1982). The New Dinosaur Dictionary. Secaucus, NJ: Citadel Press. pp. 226–228. ISBN 0-8065-0782-9.
- 1 2 Paul, G.S. (1988). "Family Spinosauridae". Predatory Dinosaurs of the World. New York: Simon & Schuster. pp. 271–274. ISBN 0-671-61946-2.
- ↑ Mortimer, M. (2007-03-25). "Comments on Therrien and Henderson's new paper". Dinosaur Mailing List. Retrieved 22 September 2010.
- ↑ Harris, J.D. (2007-03-26). "Re: Comments on Therrien and Henderson's new paper". Dinosaur Mailing List. Retrieved 22 September 2010.
- ↑ Bates, K.T.; Manning, P.L.; Hodgetts, D.; and Sellers, W.I.; Sellers, William I. (2009). Beckett, Ronald, ed. "Estimating Mass Properties of Dinosaurs Using Laser Imaging and 3D Computer Modelling". PLoS ONE. 4 (2): e4532. Bibcode:2009PLoSO...4.4532B. PMC 2639725
 . PMID 19225569. doi:10.1371/journal.pone.0004532.
. PMID 19225569. doi:10.1371/journal.pone.0004532. - 1 2 3 4 5 6 7 8 9 10 Bailey, J.B. (1997). "Neural spine elongation in dinosaurs: sailbacks or buffalo-backs?". Journal of Paleontology. 71 (6): 1124–1146. JSTOR 1306608.
- 1 2 3 4 5 6 7 Stromer, E. (1915). "Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. II. Wirbeltier-Reste der Baharije-Stufe (unterstes Cenoman). 3. Das Original des Theropoden Spinosaurus aegyptiacus nov. gen., nov. spec". Abhandlungen der Königlich Bayerischen Akademie der Wissenschaften, Mathematisch-physikalische Klasse (in German). 28 (3): 1–32.
- 1 2 3 Cuff, A. R.; Rayfield, E. J. (2013). Farke, Andrew A, ed. "Feeding Mechanics in Spinosaurid Theropods and Extant Crocodilians". PLoS ONE. 8 (5): e65295. PMC 3665537 . PMID 23724135. doi:10.1371/journal.pone.0065295.
- 1 2 3 4 5 6 7 8 Smith, J.B.; Lamanna, M.C.; Mayr, H.; and Lacovara, K.J. (2006). "New information regarding the holotype of Spinosaurus aegyptiacus Stromer, 1915". Journal of Paleontology. 80 (2): 400–406. ISSN 0022-3360. doi:10.1666/0022-3360(2006)080[0400:NIRTHO]2.0.CO;2.
- 1 2 3 4 5 6 7 8 Russell, D.A. (1996). "Isolated dinosaur bones from the Middle Cretaceous of the Tafilalt, Morocco". Bulletin du Muséum National d'Histoire Naturelle, Paris, 4e série, section C. 18 (2–3): 349–402.
- ↑ www.ted.com November 2014 Nizar Ibrahim How We Unearthed the Spinosaurus
- ↑ Stromer, E. (1934). "Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. II. Wirbeltier-Reste der Baharije-Stufe (unterstes Cenoman). 13. Dinosauria". Abhandlungen der Bayerischen Akademie der Wissenschaften Mathematisch-naturwissenschaftliche Abteilung, Neue Folge (in German). 22: 1–79.
- 1 2 3 Sereno, P.C.; Beck, A.L.; Dutheuil, D.B.; Gado, B.; Larsson, H.C.; Lyon, G.H.; Marcot, J.D.; Rauhut, O.W.M.; Sadleir, R.W.; Sidor, C.A.; Varricchio, D.J.; Wilson, G.P.; Wilson, J.A. (1998). "A long-snouted predatory dinosaur from Africa and the evolution of spinosaurids". Science. 282: 1298–1302. PMID 9812890. doi:10.1126/science.282.5392.1298.
- 1 2 Holtz, T.R., Jr.; Molnar, R.E. & Currie, P.J. (2004). "Basal Tetanurae". In Weishampel, D.B.; Dodson, P. & Osmólska, H. The Dinosauria (2nd ed.). University of California Press. pp. 71–110. ISBN 978-0-520-25408-4.
- ↑ Mahler, L. (2005). "Record of Abelisauridae (Dinosauria: Theropoda) from the Cenomanian of Morocco" (PDF). Journal of Vertebrate Paleontology. 25: 236–239. ISSN 0272-4634. doi:10.1671/0272-4634(2005)025[0236:ROADTF]2.0.CO;2.
- 1 2 Hasegawa, Y.; Tanaka, G.; Takakuwa, Y.; and Koike, S. (2010). "Fine sculptures on a tooth of Spinosaurus (Dinosauria, Theropoda) from Morocco" (PDF). Bulletin of Gunma Museum of Natural History. 14: 11–20.
- 1 2 3 4 Buffetaut, E. & Ouaja, M. (2002). "A new specimen of Spinosaurus (Dinosauria, Theropoda) from the Lower Cretaceous of Tunisia, with remarks on the evolutionary history of the Spinosauridae" (PDF). Bulletin de la Société Géologique de France. 173 (5): 415–421. doi:10.2113/173.5.415.
- 1 2 Rauhut, O.W.M. (2003). "The interrelationships and evolution of basal theropod dinosaurs". Special Papers in Palaeontology. 69: 1–213. ISBN 0-901702-79-X.
- 1 2 3 4 Taquet, P.; and Russell, D.A. (1998). "New data on spinosaurid dinosaurs from the Early Cretaceous of the Sahara" (PDF). Comptes Rendus de l'Académie des Sciences - Series IIA - Earth & Planetary Sciences. 327 (5): 347–353. Bibcode:1998CRASE.327..347T. doi:10.1016/S1251-8050(98)80054-2. Retrieved 22 September 2010.
- 1 2 3 4 Evers, S. W.; Rauhut, O. W. M.; Milner, A. C.; McFeeters, B.; Allain, R. (2015). "A reappraisal of the morphology and systematic position of the theropod dinosaur Sigilmassasaurus from the "middle" Cretaceous of Morocco". PeerJ. 3: e1323. PMC 4614847 . PMID 26500829. doi:10.7717/peerj.1323.
- ↑ "Ibrahim, N.; Sereno, P. C.; Dal Sasso, C.; Maganuco, S.; Fabbri, M.; Martill, D. M.; Zouhri, S.; Myhrvold, N.; Iurino, D. A. (2014). "Supplementary Materials for Semiaquatic Adaptations in a Giant Predatory Dinosaur". DOI: 10.1126/science.1258750".
- ↑ "Cau, Andrea (2014; online) "Spinosaurus Revolution, Episodio IV: Una soluzione a tutti gli enigmi?"http://theropoda.blogspot.com/2014/09/spinosaurus-revolution-episodio-iv-una.html".
- ↑ "Cau, Andrea (2014) "Spinosaurus Revolution, Episodio V: Sigilmassasaurus vs Spinosaurus: una battaglia tafonomica"http://theropoda.blogspot.com/2014/09/spinosaurus-revolution-episodio-v.html".
- ↑ "Mortimer, Michael. (2014) "Spinosaurus surprise"http://theropoddatabase.blogspot.com/2014/09/spinosaurus-surprise.html".
- ↑ Buffetaut, E.; Dauphin, Y.; Jaeger, J.-J.; Martin, M.; Mazin, J.-M.; and Tong, H. (1986). "Prismatic dental enamel in theropod dinosaurs". Naturwissenschaften. 73 (6): 326–327. Bibcode:1986NW.....73..326B. PMID 3748191. doi:10.1007/BF00451481.
- 1 2 Buffetaut, E. (1989). "New remains of the enigmatic dinosaur Spinosaurus from the Cretaceous of Morocco and the affinities between Spinosaurus and Baryonyx". Neues Jahrbuch für Geologie und Paläontologie, Monatshefte (2): 79–87.
- ↑ Buffetaut, E. (1992). "Remarks on the Cretaceous theropod dinosaurs Spinosaurus and Baryonyx". Neues Jahrbuch für Geologie und Paläontologie, Monatshefte (2): 88–96.
- ↑ Kellner, A.W.A.; and Mader, B.J. (1997). "Archosaur teeth from the Cretaceous of Morocco". Journal of Paleontology. 71 (3): 525–527. JSTOR 1306632.
- ↑ Benton, M.J.; Bouaziz, S.; Buffetaut, E.; Martill, D.; Ouaja, M.; Soussi, M.; and Trueman, C. (2000). "Dinosaurs and other fossil vertebrates from fluvial deposits in the Lower Cretaceous of southern Tunisia". Palaeogeography, Palaeoclimatology, Palaeoecology. 157 (3–4): 227–246. doi:10.1016/S0031-0182(99)00167-4.
- ↑ Brusatte, S. L.; and Sereno, P. C. (2007). "A new species of Carcharodontosaurus (Dinosauria: Theropoda) from the Cenomanian of Niger and a revision of the genus". Journal of Vertebrate Paleontology. 27 (4): 902–916. ISSN 0272-4634. doi:10.1671/0272-4634(2007)27[902:ANSOCD]2.0.CO;2.
- ↑ Hendrickx, C.; Mateus, O.; Buffetaut, E. (2016). "Morphofunctional Analysis of the Quadrate of Spinosauridae (Dinosauria: Theropoda) and the Presence of Spinosaurus and a Second Spinosaurine Taxon in the Cenomanian of North Africa". PLoS ONE. 11: e0144695. doi:10.1371/journal.pone.0144695.
- ↑ Weishampel, D.B.; Barrett, P.M.; Coria, R.A.; Le Loeuff, J.; Xu, X.; Zhao, X.; Sahni, A.; Gomani, E.M.P. & Noto, C.R. (2004). "Dinosaur distribution". In Weishampel, D.B.; Dodson, P. & Osmólska, H. The Dinosauria (2nd ed.). Berkeley: University of California Press. pp. 517–606. ISBN 978-0-520-25408-4.
- ↑ Allain, R.; Xaisanavong, T.; Richir, P.; Khentavong, B. (2012). "The first definitive Asian spinosaurid (Dinosauria: Theropoda) from the early cretaceous of Laos". Naturwissenschaften. 99 (5): 369–377. Bibcode:2012NW.....99..369A. PMID 22528021. doi:10.1007/s00114-012-0911-7.
- 1 2 Halstead, L.B. (1975). The Evolution and Ecology of the Dinosaurs. London: Eurobook Limited. pp. 1–116. ISBN 0-85654-018-8.
- ↑ "Gimsa, J., Sleigh, R., Gimsa, U., (2015) : "The riddle of Spinosaurus aegyptiacus ' dorsal sail". University of Rostock, Chair for Biophysics, Gertrudenstr. 11A, 18057 Rostock, Germany".
- 1 2 Charig, A.J.; Milner, A.C. (1997). "Baryonyx walkeri, a fish-eating dinosaur from the Wealden of Surrey". Bulletin of the Natural History Museum, Geology Series. 53: 11–70.
- ↑ Buffetaut, E.; Martill, D.; Escuillié, F. (2004). "Pterosaurs as part of a spinosaur diet". Nature. 430 (6995): 33. Bibcode:2004Natur.429...33B. PMID 15229562. doi:10.1038/430033a.
- 1 2 Dal Sasso, C.; Maganuco, S.; Cioffi, A. (26 May 2009). "A neurovascular cavity within the snout of the predatory dinosaur Spinosaurus" (PDF). 1st International Congress on North African Vertebrate Palaeontology. Muséum national d'Histoire naturelle. Retrieved 22 September 2010.
- 1 2 3 4 Amiot, R.; Buffetaut, E.; Lécuyer, C.; Wang, X.; Boudad, L.; Ding, Z.; Fourel, F.; Hutt, S.; Martineau, F.; Medeiros, A.; Mo, J.; Simon, L.; Suteethorn, V.; Sweetman, S.; Tong, H.; Zhang, F.; Zhou, Z. (2010). "Oxygen isotope evidence for semi-aquatic habits among spinosaurid theropods". Geology. 38 (2): 139–142. doi:10.1130/G30402.1.
- 1 2 3 Glut, D.F. (2000). "Spinosaurus". Dinosaurs: The Encyclopedia. 1st Supplement. Jefferson, North Carolina: McFarland & Company, Inc. pp. 329–333. ISBN 0-7864-0591-0.
- ↑ Carpenter, K. (2002). "Forelimb biomechanics of nonavian theropod dinosaurs in predation". Senckenbergiana Lethaea. 82 (1): 59–76. doi:10.1007/BF03043773.
- ↑ Milner, A.R.C.; Harris, J.D.; Lockley, M.G.; Kirkland, J.I.; Matthews, N.A. (2009). Harpending, Henry, ed. "Bird-like anatomy, posture, and behavior revealed by an Early Jurassic theropod dinosaur resting trace". PLoS ONE. 4 (3): e4591. Bibcode:2009PLoSO...4.4591M. PMC 2645690 . PMID 19259260. doi:10.1371/journal.pone.0004591.
- ↑ Ibrahim, N.; Sereno, P. C.; Dal Sasso, C.; Maganuco, S.; Fabbri, M.; Martill, D. M.; Zouhri, S.; Myhrvold, N.; Iurino, D. A. (2014). "Semiaquatic adaptations in a giant predatory dinosaur". Science. 345 (6204): 1613–1616. PMID 25213375. doi:10.1126/science.1258750.
- ↑ Witze, A. (2014). "Swimming dinosaur found in Morocco". Nature. doi:10.1038/nature.2014.15901.
- ↑ Scott Hartman (12 September 2014). "There's something fishy about Spinosaurus". skeletaldrawing.com. Retrieved 20 September 2014.
- ↑ Mark Witton (22 September 2014). "The Spinosaurus hindlimb controversy: a detailed response from the authors". markwitton-com.blogspot.de. Retrieved 22 September 2014.
- ↑ Gradstein, F.M.; Ogg, J.G.; Smith, A.G., eds. (2004). A Geologic Time Scale 2004. Cambridge and New York: Cambridge University Press. p. 380. ISBN 0-521-78673-8.
- ↑ Sereno, PC; Dutheil, DB; Iarochene, M; Larsson, HCE; Lyon, GH; Magwene, PM; Sidor, CA; Varricchio, DJ; Wilson, JA (1996). "Predatory dinosaurs from the Sahara and Late Cretaceous faunal differentiation". Science. 272: 986–991. PMID 8662584. doi:10.1126/science.272.5264.986.
- ↑ Smith, J.B.; Lamanna, M.C.; Lacovara, K.J.; Dodson, P.; Smith, J.R.; Poole, J.C.; Giegengack, R.; Attia, Y. (2001). "A giant sauropod dinosaur from an Upper Cretaceous mangrove deposit in Egypt". Science. 292 (5522): 1704–1706. Bibcode:2001Sci...292.1704S. PMID 11387472. doi:10.1126/science.1060561.
- ↑ Naish, Darren (2012). Planet Dinosaur : The Next Generation of Killer Giants. Firefly Books. pp. 80–85. ISBN 978-1-77085-049-1.
- ↑ Elley, Derek (July 17, 2001). "Jurassic Park III". Variety. Retrieved July 9, 2007.
- ↑ Portman, J. (11 July 2001). "Spinosaurus makes T. Rex look like a pussycat: When it comes to Jurassic Park III, size does matter". Ottawa Citizen.
- ↑ Chandler, G. (August 2001). "A bite-size guide to the dinosaurs of the new movie Jurassic Park III". National Geographic World.
- ↑ Romano, Nick (June 2015). "Jurassic World Snuck in A Sweet Nod to Jurassic Park 3". CinemaBlend.
- ↑ Lapparent, A.F. de; and Lavocat, R. (1955). "Dinosauriens." In: Piveteau, J., editor. Traité de Paléontologie. Tome V. La Sortie des Eaux. Naissance de la Tétrapodie. L'Exubérance de la Vie Végétative. La Conquête de l'Air. Amphibiens. Reptiles. Oiseaux. Paris: Masson et Cie, pp. 785-962.
- ↑ Stromer, E. (1936). "Ergebnisse der Forschungsreisen Prof. E. Stromers in den Wüsten Ägyptens. VII. Baharije-Kessel und -Stufe mit deren Fauna und Flora. Eine ergänzende Zusammenfassung". Abhandlungen der Bayerischen Akademie der Wissenschaften, Mathematisch-naturwissenschaftliche Abteilung, Neue Folge (in German). 33: 1–102.
- ↑ Khatri, V.S. (9 June 2006). "From the past". The Hindu. Retrieved 12 September 2010.
- ↑ Glut, D.F.; and Brett-Surman, M.K. (2000). "Dinosaurs and the media" (PDF). In Farlow, J.O.; and Walters, R.H. The Complete Dinosaur. Bloomington, IN: Indiana University Press. pp. 673–706. ISBN 0-253-21313-4. Retrieved 12 September 2010.
Further reading
- Glut, D.F. "In search of Spinosaurus." In: Jurassic classics: a collection of saurian essays and Mesozoic musings, pp. 77–85. Jefferson, NC: McFarland, 2001. ISBN 0-7864-0961-4.
- Nothdurft, W.; and Smith, J. The Lost Dinosaurs of Egypt. New York: Random House, 2002. ISBN 0-375-50795-7.
- A Tribute to Ernst Stromer: Hundred Years of the Discovery of Spinosaurus aegypticus: Saubhik Ghosh
External links
| Wikimedia Commons has media related to Spinosaurus. |
| Wikiquote has quotations related to: Spinosaurus |
| Wikispecies has information related to: Spinosaurus |
- Hartman, Scott. Spinosaur Comparison. SkeletalDrawing.com, 2006.
- Lloyd, Robin. The Biggest Carnivore: Dinosaur History Rewritten. LiveScience, 1 March 2006.
- Mortimer, Mickey. Spinosaurus Stromer, 1915. (List of specimens from The Theropod Database.)
- Natural History Museum. Dino Directory: Spinosaurus.
- Prendergast, John. Dinosaurs Lost and Found. The Pennsylvania Gazette, the Alumni Magazine of the University of Pennsylvania, July/Aug 2001.
- The Top 8 Paleontology Stories of 2006: 40 New Dinosaur King Rears Its Head. Discover Magazine, 29 December 2006.
| |||||||||||||||||||
| Piatnitzkysauridae | | ||||||||||||||||||
| Megalosauria | |||||||||||||||||||