SAMHD1
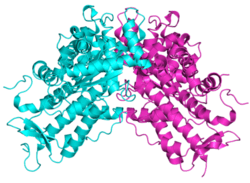




SAM domain and HD domain-containing protein 1 is a protein that in humans is encoded by the SAMHD1 gene. SAMHD1 is a cellular enzyme, responsible for blocking replication of HIV in dendritic cells,[5] macrophages[6] and monocytes.[7] It is an enzyme that exhibits phosphohydrolase activity,[8][9] converting deoxynucleoside triphosphates (dNTPs) to inorganic phosphate (iPPP) and a 2'-deoxynucleoside (i.e. deoxynucleosides without a phosphate group). In doing so, SAMHD1 depletes the pool of dNTPs available to a reverse transcriptase for viral cDNA synthesis and thus prevents viral replication.[10] SAMHD1 has also shown nuclease activity.[11] Although a ribonuclease activity was described to be required for HIV-1 restriction,[12] recent data confirmed that SAMHD1-mediated HIV-1 restriction in cells does not involve ribonuclease activity.[13]
Nomenclature
The SAMHD1 protein is also known as:
- AGS5: Aicardi- Goutières syndrome type 5[14]
- DCIP: Dendritic cell-derived IFNG-induced protein2[15]
- Mg11: Interferon-gamma-inducible protein[15]
- HDDC1: HD domain containing 1
- MOP-5: Monocyte protein 5
- SAMH1_HUMAN
- SBBI88
- CHBL2
Gene
The gene encoding human SAMHD1 was originally identified in a human dendritic cell cDNA library as an orthologue of a mouse gene IFN-γ-induced gene Mg11.[15] The SAMHD1 gene is located on chromosome 20. SAMHD1 spans 59,532 bp of genomic sequence (chromosome 20:34,954,059–35,013,590) in 16 exons and encodes a 626 amino-acid (aa) protein with a molecular weight of 72.2 kDa.[16][17] SAMHD1 expressed in both cycling and noncycling cells, but the antiviral activity of SAMHD1 is limited to noncycling cells.[18]
Structure
The SAMHD1 is 626 amino acids (aa) long and has 2 domains:
a. Sterile Alpha Motif (SAM) domain: residues 45 – 110 aa.[19][20] In general, SAM domains are known to function as protein–protein and protein–nucleic acid interactions in organisms from yeast to humans, docking sites for kinases, signal transduction and regulation of transcription.[21][22]
b. Histidine- Aspartic (HD) domain-containing protein 1: residues 164 – 319 aa.[19][20] HD domains proteins are characterized by a doublet of histidine and aspartic acid catalytic residues, and have been shown to possess putative nuclease, dGTP triphosphatase, phosphatase or phosphodiesterase activities.[21][23]
A crystal structure of a SAMHD1 fragment comprising catalytic core reveals that the protein is dimeric.[9] Also studies have shown that SAMHD1 oligomerizes and forms tetramers.[24] SAMHD1 is phosphorylated on residue T592 in cycling cells but that this phosphorylation is lost when cells are in a noncycling state. [25]
Function
Mutations in SAMHD1 are found in Aicardi–Goutières syndrome (AGS), “a hereditary autoimmune encephalopathy that is characterized by aberrant production of type I interferon (IFN) and symptoms mimicking congenital viral infection”.[20] Monocytes isolated from individuals with AGS are highly susceptible to HIV-1.[7]
SAMHD1 was identified as a host protein that is bound and blocked by lentiviral protein, Vpx. Vpx promotes macrophage and DC infection by targeting SAMHD1.[26]
The human SAMHD1 protein has dNTP triphosphatase activity, specifically dGTP-stimulated dNTP triphosphohydrolase activity, and nuclease activity against single-stranded DNA and RNA which is associated with its HD domain.[11][27] Other studies demonstrated that silencing SAMHD1 enhanced HIV-1 and SIV Δvpx infection of myeloid cells, also enhances HIV-1 infection of resting CD4+ T cells.[18][27]
Role in disease
Aicardi-Goutieres syndrome
16 mutations in the SAMHD1 gene have been identified in patients with Aicardi-Goutieres syndrome. Mutations result in a SAMHD1 less functional protein. However, it is not known how this protein dysfunction leads to immune system abnormalities, inflammatory damage to the brain and skin, and other characteristics of this syndrome.[7][20]
Restriction viral infection
SAMHD1 was identified as the cellular protein responsible of the reverse transcription block to HIV-1 infection observed in myeloid cells as well as in quiescent CD4+ T cells. SAMHD1 inhibits HIV-1 infection in myeloid cells by limiting the intracellular pool of dNTPs.[26] The dNTP triphosphohydrolase activity of SAMHD1 has been proposed to reduce the intracellular dNTP level, restricting HIV-1 replication and preventing activation of the immune system, a nuclease activity against single-stranded (ss)DNAs and RNAs, as well as against RNA in DNA/RNA hybrids.[21][26] Retroviral restriction ability of SAMHD1 is regulated by phosphorylation, for this purpose SAMHD1 associates with the cyclin A2/CDK1 complex that mediates its phosphorylation at threonine 592. Phosphorylated SAMHD1 has been observed to have minimal to no activity in cycling cells. Conversely, unphosphorylated SAMHD1 in non-cycling cells have potent restriction activity.[25]
Expression modulation and antimetabolite degradation in cancer cells
SAMHD1 protein expression can be influenced at four levels in cancer cells. First, mutations in the SAMHD1 gene can prevent SAMHD1 mRNA generation or a functional protein after translations.[28] Second, promoter methylation can prevent SAMHD1 mRNA transcription.[29][30] Third, miRNA-155 and miRNA-181a can prevent the translation, thus preventing protein production.[31][32] Finally, SAMHD1 degradation occurs during the S phase of the cell cycle.[33] Non-adherent tumor cell lines – B cells, T cells and myeloid cells can be rapidly dividing cells, have low to no detectable levels of SAMHD1 protein, as compared to adherent cells. Regulation of dNTP concentration by SAMHD1 in cancer cells might be an important mechanism with therapeutic implications.[34] Antimetabolites are anticancer nucleosides, nucleotides, and base analogs used as anticancer agents to promote cell death by several different mechanisms of action.[35] For some of these antimetabolites, the intracellular triphosphate form of the analog is the active compound.[35] SAMHD1 has been shown to be able to hydrolyze arabinose 5’-triphosphates.[36] SAMHD1 has been shown to be a biomarker and influence arabinose C (ara-C; cytarabine) responsiveness.[37] Viral protein x (Vpx) has been proposed to be potential therapy to improve cytarabine therapy for hematological malignancies.[38]
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000101347 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000027639 - Ensembl, May 2017
- ↑ "Human PubMed Reference:".
- ↑ "Mouse PubMed Reference:".
- ↑ Laguette N, Sobhian B, Casartelli N, Ringeard M, Chable-Bessia C, Ségéral E, Yatim A, Emiliani S, Schwartz O, Benkirane M (May 2011). "SAMHD1 is the dendritic- and myeloid-cell-specific HIV-1 restriction factor counteracted by Vpx". Nature. 474 (7353): 654–7. PMC 3595993
 . PMID 21613998. doi:10.1038/nature10117.
. PMID 21613998. doi:10.1038/nature10117. - ↑ Hrecka K, Hao C, Gierszewska M, Swanson SK, Kesik-Brodacka M, Srivastava S, Florens L, Washburn MP, Skowronski J (June 2011). "Vpx relieves inhibition of HIV-1 infection of macrophages mediated by the SAMHD1 protein". Nature. 474 (7353): 658–61. PMC 3179858 . PMID 21720370. doi:10.1038/nature10195.
- 1 2 3 Berger A, Sommer AF, Zwarg J, Hamdorf M, Welzel K, Esly N, Panitz S, Reuter A, Ramos I, Jatiani A, Mulder LC, Fernandez-Sesma A, Rutsch F, Simon V, König R, Flory E (December 2011). "SAMHD1-deficient CD14+ cells from individuals with Aicardi-Goutières syndrome are highly susceptible to HIV-1 infection". PLoS Pathogens. 7 (12): e1002425. PMC 3234228 . PMID 22174685. doi:10.1371/journal.ppat.1002425.
- ↑ Powell RD, Holland PJ, Hollis T, Perrino FW (December 2011). "Aicardi-Goutieres syndrome gene and HIV-1 restriction factor SAMHD1 is a dGTP-regulated deoxynucleotide triphosphohydrolase". The Journal of Biological Chemistry. 286 (51): 43596–600. PMC 3243528 . PMID 22069334. doi:10.1074/jbc.C111.317628.
- 1 2 Goldstone DC, Ennis-Adeniran V, Hedden JJ, Groom HC, Rice GI, Christodoulou E, Walker PA, Kelly G, Haire LF, Yap MW, de Carvalho LP, Stoye JP, Crow YJ, Taylor IA, Webb M (November 2011). "HIV-1 restriction factor SAMHD1 is a deoxynucleoside triphosphate triphosphohydrolase". Nature. 480 (7377): 379–82. PMID 22056990. doi:10.1038/nature10623.
- ↑ Lahouassa H, Daddacha W, Hofmann H, Ayinde D, Logue EC, Dragin L, Bloch N, Maudet C, Bertrand M, Gramberg T, Pancino G, Priet S, Canard B, Laguette N, Benkirane M, Transy C, Landau NR, Kim B, Margottin-Goguet F (February 2012). "SAMHD1 restricts the replication of human immunodeficiency virus type 1 by depleting the intracellular pool of deoxynucleoside triphosphates". Nature Immunology. 13 (3): 223–8. PMC 3771401 . PMID 22327569. doi:10.1038/ni.2236.
- 1 2 Beloglazova N, Flick R, Tchigvintsev A, Brown G, Popovic A, Nocek B, Yakunin AF. “Nuclease activity of the human SAMHD1 protein implicated in the Aicardi-Goutieres syndrome and HIV-1 restriction”. Beloglazova N, Flick R, Tchigvintsev A, Brown G, Popovic A, Nocek B, Yakunin AF (March 2013). "Nuclease activity of the human SAMHD1 protein implicated in the Aicardi-Goutieres syndrome and HIV-1 restriction". The Journal of Biological Chemistry. 288 (12): 8101–10. PMC 3605629 . PMID 23364794. doi:10.1074/jbc.M112.431148.
- ↑ Ryoo J, Choi J, Oh C, Kim S, Seo M, Kim SY, Seo D, Kim J, White TE, Brandariz-Nuñez A, Diaz-Griffero F, Yun CH, Hollenbaugh JA, Kim B, Baek D, Ahn K (August 2014). "The ribonuclease activity of SAMHD1 is required for HIV-1 restriction". Nature Medicine. 20 (8): 936–41. PMC 4318684 . PMID 25038827. doi:10.1038/nm.3626.
- ↑ Antonucci JM, St Gelais C, de Silva S, Yount JS, Tang C, Ji X, Shepard C, Xiong Y, Kim B, Wu L (October 2016). "SAMHD1-mediated HIV-1 restriction in cells does not involve ribonuclease activity". Nature Medicine. 22 (10): 1072–1074. PMC 5069697 . PMID 27711056. doi:10.1038/nm.4163.
- ↑ Chahwan C, Chahwan R (May 2012). "Aicardi-Goutieres syndrome: from patients to genes and beyond". Clinical Genetics. 81 (5): 413–20. PMID 22149989. doi:10.1111/j.1399-0004.2011.01825.x.
- 1 2 3 Li N, Zhang W, Cao X (November 2000). "Identification of human homologue of mouse IFN-gamma induced protein from human dendritic cells". Immunology Letters. 74 (3): 221–4. PMID 11064105. doi:10.1016/S0165-2478(00)00276-5.
- ↑ Rice GI, Bond J, Asipu A, Brunette RL, Manfield IW, Carr IM, et al. (July 2009). "Mutations involved in Aicardi-Goutières syndrome implicate SAMHD1 as regulator of the innate immune response". Nature Genetics. 41 (7): 829–32. PMC 4154505 . PMID 19525956. doi:10.1038/ng.373.
- ↑ Welbourn S, Miyagi E, White TE, Diaz-Griffero F, Strebel K (October 2012). "Identification and characterization of naturally occurring splice variants of SAMHD1". Retrovirology. 9: 86. PMC 3503569 . PMID 23092512. doi:10.1186/1742-4690-9-86.
- 1 2 Baldauf HM, Pan X, Erikson E, Schmidt S, Daddacha W, Burggraf M, Schenkova K, Ambiel I, Wabnitz G, Gramberg T, Panitz S, Flory E, Landau NR, Sertel S, Rutsch F, Lasitschka F, Kim B, König R, Fackler OT, Keppler OT (November 2012). "SAMHD1 restricts HIV-1 infection in resting CD4(+) T cells". Nature Medicine. 18 (11): 1682–7. PMC 3828732 . PMID 22972397. doi:10.1038/nm.2964.
- 1 2 White TE, Brandariz-Nuñez A, Valle-Casuso JC, Amie S, Nguyen L, Kim B, Brojatsch J, Diaz-Griffero F (February 2013). "Contribution of SAM and HD domains to retroviral restriction mediated by human SAMHD1". Virology. 436 (1): 81–90. PMC 3767443 . PMID 23158101. doi:10.1016/j.virol.2012.10.029.
- 1 2 3 4 Crow YJ, Rehwinkel J (October 2009). "Aicardi-Goutieres syndrome and related phenotypes: linking nucleic acid metabolism with autoimmunity". Human Molecular Genetics. 18 (R2): R130–6. PMC 2758706 . PMID 19808788. doi:10.1093/hmg/ddp293.
- 1 2 3 Ayinde D, Casartelli N, Schwartz O (October 2012). "Restricting HIV the SAMHD1 way: through nucleotide starvation". Nature Reviews. Microbiology. 10 (10): 675–80. PMID 22926205. doi:10.1038/nrmicro2862.
- ↑ Kim CA, Bowie JU (December 2003). "SAM domains: uniform structure, diversity of function". Trends in Biochemical Sciences. 28 (12): 625–8. PMID 14659692. doi:10.1016/j.tibs.2003.11.001.
- ↑ Aravind L, Koonin EV (December 1998). "The HD domain defines a new superfamily of metal-dependent phosphohydrolases". Trends in Biochemical Sciences. 23 (12): 469–72. PMID 9868367. doi:10.1016/s0968-0004(98)01293-6.
- ↑ Yan J, Kaur S, DeLucia M, Hao C, Mehrens J, Wang C, Golczak M, Palczewski K, Gronenborn AM, Ahn J, Skowronski J (April 2013). "Tetramerization of SAMHD1 is required for biological activity and inhibition of HIV infection". The Journal of Biological Chemistry. 288 (15): 10406–17. PMC 3624423 . PMID 23426366. doi:10.1074/jbc.M112.443796.
- 1 2 White TE, Brandariz-Nuñez A, Valle-Casuso JC, Amie S, Nguyen LA, Kim B, Tuzova M, Diaz-Griffero F (April 2013). "The retroviral restriction ability of SAMHD1, but not its deoxynucleotide triphosphohydrolase activity, is regulated by phosphorylation". Cell Host & Microbe. 13 (4): 441–51. PMC 3864637 . PMID 23601106. doi:10.1016/j.chom.2013.03.005.
- 1 2 3 Mashiba M, Collins KL (January 2013). "Molecular mechanisms of HIV immune evasion of the innate immune response in myeloid cells". Viruses. 5 (1): 1–14. PMC 3564108 . PMID 23344558. doi:10.3390/v5010001.
- 1 2 Descours B, Cribier A, Chable-Bessia C, Ayinde D, Rice G, Crow Y, Yatim A, Schwartz O, Laguette N, Benkirane M (October 2012). "SAMHD1 restricts HIV-1 reverse transcription in quiescent CD4(+) T-cells". Retrovirology. 9: 87. PMC 3494655 . PMID 23092122. doi:10.1186/1742-4690-9-87.
- ↑ Clifford R, Louis T, Robbe P, Ackroyd S, Burns A, Timbs AT, et al. (February 2014). "SAMHD1 is mutated recurrently in chronic lymphocytic leukemia and is involved in response to DNA damage". Blood. 123 (7): 1021–31. PMC 3924925 . PMID 24335234. doi:10.1182/blood-2013-04-490847.
- ↑ Wang JL, Lu FZ, Shen XY, Wu Y, Zhao LT (December 2014). "SAMHD1 is down regulated in lung cancer by methylation and inhibits tumor cell proliferation". Biochemical and Biophysical Research Communications. 455 (3–4): 229–33. PMID 25449277. doi:10.1016/j.bbrc.2014.10.153.
- ↑ de Silva S, Wang F, Hake TS, Porcu P, Wong HK, Wu L (February 2014). "Downregulation of SAMHD1 expression correlates with promoter DNA methylation in Sézary syndrome patients". The Journal of Investigative Dermatology. 134 (2): 562–5. PMC 3844041 . PMID 23884314. doi:10.1038/jid.2013.311.
- ↑ Kohnken R, Kodigepalli KM, Mishra A, Porcu P, Wu L (January 2017). "MicroRNA-181 contributes to downregulation of SAMHD1 expression in CD4+ T-cells derived from Sèzary syndrome patients". Leukemia Research. 52: 58–66. PMC 5195900 . PMID 27889686. doi:10.1016/j.leukres.2016.11.010.
- ↑ Pilakka-Kanthikeel S, Raymond A, Atluri VS, Sagar V, Saxena SK, Diaz P, Chevelon S, Concepcion M, Nair M (April 2015). "Sterile alpha motif and histidine/aspartic acid domain-containing protein 1 (SAMHD1)-facilitated HIV restriction in astrocytes is regulated by miRNA-181a". Journal of Neuroinflammation. 12: 66. PMC 4410490 . PMID 25890101. doi:10.1186/s12974-015-0285-9.
- ↑ Kretschmer S, Wolf C, König N, Staroske W, Guck J, Häusler M, Luksch H, Nguyen LA, Kim B, Alexopoulou D, Dahl A, Rapp A, Cardoso MC, Shevchenko A, Lee-Kirsch MA (March 2015). "SAMHD1 prevents autoimmunity by maintaining genome stability". Annals of the Rheumatic Diseases. 74 (3): e17. PMC 4345975 . PMID 24445253. doi:10.1136/annrheumdis-2013-204845.
- ↑ Kohnken R, Kodigepalli KM, Wu L (September 2015). "Regulation of deoxynucleotide metabolism in cancer: novel mechanisms and therapeutic implications". Molecular Cancer. 14: 176. PMC 4587406 . PMID 26416562. doi:10.1186/s12943-015-0446-6.
- 1 2 Shelton J, Lu X, Hollenbaugh JA, Cho JH, Amblard F, Schinazi RF (December 2016). "Metabolism, Biochemical Actions, and Chemical Synthesis of Anticancer Nucleosides, Nucleotides, and Base Analogs". Chemical Reviews. 116 (23): 14379–14455. PMID 27960273. doi:10.1021/acs.chemrev.6b00209.
- ↑ Hollenbaugh JA, Shelton J, Tao S, Amiralaei S, Liu P, Lu X, Goetze RW, Zhou L, Nettles JH, Schinazi RF, Kim B (Jan 2017). "Substrates and Inhibitors of SAMHD1". PloS One. 12 (1): e0169052. PMC 5207538 . PMID 28046007. doi:10.1371/journal.pone.0169052.
- ↑ Schneider C, Oellerich T, Baldauf HM, Schwarz SM, Thomas D, Flick R, et al. (February 2017). "SAMHD1 is a biomarker for cytarabine response and a therapeutic target in acute myeloid leukemia". Nature Medicine. 23 (2): 250–255. PMID 27991919. doi:10.1038/nm.4255.
- ↑ Herold N, Rudd SG, Ljungblad L, Sanjiv K, Myrberg IH, Paulin CB, et al. (February 2017). "Targeting SAMHD1 with the Vpx protein to improve cytarabine therapy for hematological malignancies". Nature Medicine. 23 (2): 256–263. PMID 28067901. doi:10.1038/nm.4265.
Further reading
- Laguette N, Benkirane M (January 2012). "How SAMHD1 changes our view of viral restriction". Trends in Immunology. 33 (1): 26–33. PMC 3595995 . PMID 22177690. doi:10.1016/j.it.2011.11.002.
- Olsen JV, Blagoev B, Gnad F, Macek B, Kumar C, Mortensen P, Mann M (November 2006). "Global, in vivo, and site-specific phosphorylation dynamics in signaling networks". Cell. 127 (3): 635–48. PMID 17081983. doi:10.1016/j.cell.2006.09.026.
External links
- GeneReviews/NCBI/NIH/UW entry on Aicardi-Goutières Syndrome
- OMIM entries on Aicardi-Goutieres syndrome