Radial unit hypothesis
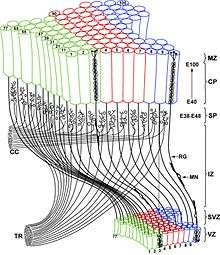
The Radial Unit Hypothesis (RUH) is a conceptual theory of cerebral cortex development, first described by Pasko Rakic. The RUH states that the cerebral cortex develops during embryogenesis as an array of interacting cortical columns, or 'radial units', each of which originates from a transient stem cell layer called the ventricular zone, which contains neural stem cells known as radial glial cells.[1][2] The reiterative nature of the cerebral cortex, in the sense that it is a vast array of repeating functional circuits, led to the idea that cortical evolution is governed by mechanisms regulating the addition of cortical columns, enabling additional functional areas to become specialized and incorporated into the brain. The addition of new radial units is thought to depend on control of the cell cycle (proliferation) of cortical stem cells lining the ventricular system in the ventricular zone and subventricular zone.[3]
Intimately related to the RUH is the 'protomap' hypothesis, which states that the primordial identity of each functional area of the cerebral cortex is encoded within the cortical stem cells prior to the formation of the cortical layers.[1] Within each developing radial unit, the process of neurogenesis gives rise to post-mitotic (non-dividing) cortical neurons, which begin the process of radial neuronal migration from the ventricular zone and adjacent subventricular zone to form the cortical plate in the classic 'inside-out' manner beginning with the deep cortical layers.[4][5] Once their final destination is achieved, cortical neurons begin to form circuits with other cortical and subcortical neurons, often taking on a columnar shape following the radial migration route.[6] Some localized lateral dispersion takes place during cortical column development in the mouse, but the degree of dispersion is molecularly regulated and indeed could vary across species.[7]
Together, the RUH and protomap hypothesis represent two core principles of early cerebral cortex development.[3] After neurons arrive in the cortical plate, other processes—especially activity-dependent processes—govern the maturation of cortical circuitry.[8]
See also
- Neural stem cell
- Stem cell
- Ventricular zone
- Subventricular zone
- Neurogenesis
- Cellular differentiation
- Cortical patterning
- Protomap
- Cortical column
References
- 1 2 Rakic, P (8 July 1988). "Specification of cerebral cortical areas.". Science. 241 (4862): 170–6. PMID 3291116. doi:10.1126/science.3291116.
- ↑ Rakic, P (September 1995). "A small step for the cell, a giant leap for mankind: a hypothesis of neocortical expansion during evolution.". Trends in Neurosciences. 18 (9): 383–8. PMID 7482803. doi:10.1016/0166-2236(95)93934-p.
- 1 2 Rakic, P (October 2009). "Evolution of the neocortex: a perspective from developmental biology.". Nature Reviews. Neuroscience. 10 (10): 724–35. PMC 2913577
 . PMID 19763105. doi:10.1038/nrn2719.
. PMID 19763105. doi:10.1038/nrn2719. - ↑ Rakic, P (May 1972). "Mode of cell migration to the superficial layers of fetal monkey neocortex.". The Journal of Comparative Neurology. 145 (1): 61–83. PMID 4624784. doi:10.1002/cne.901450105.
- ↑ Rakic, P (1 February 1974). "Neurons in rhesus monkey visual cortex: systematic relation between time of origin and eventual disposition.". Science. 183 (4123): 425–7. PMID 4203022. doi:10.1126/science.183.4123.425.
- ↑ Yuste, R; Peinado, A; Katz, LC (31 July 1992). "Neuronal domains in developing neocortex.". Science. 257 (5070): 665–9. PMID 1496379. doi:10.1126/science.1496379.
- ↑ Torii, M; Hashimoto-Torii, K; Levitt, P; Rakic, P (24 September 2009). "Integration of neuronal clones in the radial cortical columns by EphA and ephrin-A signalling.". Nature. 461 (7263): 524–8. PMC 2874978 . PMID 19759535. doi:10.1038/nature08362.
- ↑ Ackman, JB; Burbridge, TJ; Crair, MC (11 October 2012). "Retinal waves coordinate patterned activity throughout the developing visual system.". Nature. 490 (7419): 219–25. PMC 3962269 . PMID 23060192. doi:10.1038/nature11529.