Haplogroup R1b
| Haplogroup R1b | |
|---|---|
.PNG) | |
| Possible place of origin | Western Asia[1] |
| Ancestor | R1 |
| Descendants |
One immediate descendant:
|
| Defining mutations | M343 |
Haplogroup R1b (R-M343), also known as Hg1 and Eu18, is a human Y-chromosome haplogroup.
It is the most frequently occurring paternal lineage in Western Europe, as well as some parts of Russia (e.g. the Bashkir minority) and Central Africa (e.g. Chad and Cameroon). The clade is also present at lower frequencies throughout Eastern Europe, Western Asia, as well as parts of North Africa and Central Asia.
R1b also reaches high frequencies in the Americas and Australasia, due largely to immigration from Western Europe. There is an ongoing debate regarding the origins of R1b subclades found at significant levels among some indigenous peoples of the Americas, such as speakers of Algic languages in central Canada.
R1b has one primary branch, R1b1 (L278), which in turn has two primary branches: R1b1a (L754) and R1b1b (PH155). R1b1a is found mostly in Western Europe, although the Fula and Chadic-speaking peoples of Africa are dominated by R1b1a2 (PF6279/V88).
R1b1b (PH155) is so rare and widely dispersed that it is difficult to draw any conclusions about its origins. It has been found in Bahrain, Bhutan, Tajikistan, Turkey and Western China.
Western Europe is dominated by the downstream subclades of R1b1a – especially R1b1a1a2 (R-M269; known previously as R1b1a2).
The oldest human remains found to carry with R1b so far are:
- an individual from an Epigravettian culture context in Italy (Villabruna) who lived circa 12,000 BC and reportedly belonged to R1b1a (L754), and
- the 7,000-year-old remains of a hunter-gatherer, belonging to the Samara culture of the Volga River area, who carried R1b1* (R-L278*).
Origin and dispersal
R1b is a subclade within the "macro-haplogroup" Haplogroup K (K-M9), which is one of the predominant groupings of all the rest of human male lines outside of Africa. K* is believed to have originated in Asia (as is the case with an even earlier ancestral haplogroup, F (F-M89). Karafet T. et al. (2014) "rapid diversification process of K-M526 likely occurred in Southeast Asia, with subsequent westward expansions of the ancestors of haplogroups R and Q." [2]
Three autosomal genetic studies in 2015 gave support to the Kurgan hypothesis of Gimbutas regarding the proto-Indo-European homeland. According to those studies, haplogroups R1b and R1a, now the most common in Europe (R1a is also common in South Asia) would have expanded from the Russian steppes, along with the Indo European languages; they also detected an autosomal component present in modern Europeans which was not present in Neolithic Europeans, which would have been introduced with paternal lineages R1b and R1a, as well as Indo European Languages.[3][4][5]
The point of origin of R1b is thought to lie in Eurasia, most likely in Western Asia.[6] T. Karafet et al. (2008) estimated the age of R1, the parent of R1b, as 18,500 years before present.[7]
Early research into the origins of R1b focused on Europe. In 2000, Ornella Semino and colleagues argued that R1b had been in Europe before the end of the Ice Age, and had spread north from an Iberian refuge after the Last Glacial Maximum.[8] Age estimates of R1b in Europe have steadily decreased in more recent studies, at least concerning the majority of R1b, with more recent studies suggesting a Neolithic age or younger.[6][9][10][11] On the other hand, Morelli et al. have recently (in 2010) attempted to defend a Palaeolithic origin for R1b1b2.[12] Irrespective of microsatellite coalescence calculations, Chikhi et al. pointed out that the timing of molecular divergences does not coincide with population splits; the TMRCA of haplogroup R1b (whether in the Palaeolithic or Neolithic) dates to its point of origin somewhere in Eurasia, and not its arrival in western Europe.[13] Summing up, Michael R. Maglio argues that the closest branch of R1b is from Iberia and its small subclades found in West Asia, the Near East and Africa are examples of back migration, and not of its origin.[14]
However, as Barbara Arredi and colleagues were the first to point out, the distribution of R1b microsatellite variance in Europe forms a cline from east to west, which is more consistent with an entry into Europe from Western Asia with the spread of farming.[11] A 2009 paper by Chiaroni et al. added to this perspective by using R1b as an example of a wave haplogroup distribution, in this case from east to west.[15] The proposal of a southeastern origin of R1b were supported by three detailed studies based on large datasets published in 2010. These detected that the earliest subclades of R1b are found in western Asia and the most recent in western Europe.[6][9][16]
While age estimates in these articles are all more recent than the Last Glacial Maximum, all mention the Neolithic, when farming was introduced to Europe from the Middle East as a possible candidate period. Myres et al. (August 2010), and Cruciani et al. (August 2010) both remained undecided on the exact dating of the migration or migrations responsible for this distribution, not ruling out migrations as early as the Mesolithic or as late as the Hallstatt culture but more probably Late Neolithic.[6] They noted that direct evidence from ancient DNA may be needed to resolve these gene flows.[6] Lee et al. (May 2012) analysed the ancient DNA of human remains from the Late Neolithic Beaker culture site of Kromsdorf, Germany identifying two males as belonging to the Y haplogroup R1b.[17] Analysis of ancient Y DNA from the remains of populations derived from early Neolithic Central and North European Linear Pottery culture settlements have not yet found males belonging to haplogroup R1b.[18][19]
The suggestion has also been made that the spread of R1b in Western Europe may coincide with the spread of the Centum branch of the Indo-European languages during the early Bronze Age.[20]
One of the highest levels of R1b is found among the Basques, who speak a non- Indo-European language isolate; which contradicts an exclusively or predominantly Indo-European source. One hypothesis about the case of the Basques is that a male-dominated Indo-European-speaking people invaded and conquered the Basque region, and then, having brought no or few women with them, married local women, possibly from a matrilineal society. The women then raised the children that resulted to speak their own language and with their cultural practices, rather than those of the fathers'. This possible explanation further addresses the point that while other high-R1b regions in Western Europe (such as the British Isles and southern Germany) show disproportionately high incidences of MtDNA haplogroups that correspond to a Pontic Steppes origin (specifically MtDNA Haplogroups I, U2, U3, U4, and W), the Basque region does not. In fact, the Basque region displays virtually no MtDNA for which Pontic Steppes origin could be claimed.[20]
In the 2016 Nature article 'The genetic history of Ice Age Europe', an individual from the Epigravettian cultural context in Italy (Villabruna) is mentioned, who lived circa 12,000 BC and reportedly belonged to Y-DNA group R1b1a (L754).
Structure
External phylogeny
R1b is a part of the broader Haplogroup K-M9 and its linear descendants K2, K2b and P, which is also known as K2b2.
Karafet et al. (2014) suggest that a "rapid diversification process of K-M526 likely occurred in Southeast Asia, with subsequent westward expansions of the ancestors of haplogroups R and Q." [2]
Internal structure
Names such as R1b, R1b1 and so on are phylogenetic (i.e. "family tree") names which make clear their place within the branching of haplogroups, or the phylogenetic tree. An alternative way of naming the same haplogroups and subclades refers to their defining SNP mutations: for example, R-M343 is equivalent to R1b.[21] Phylogenetic names change with new discoveries and SNP-based names are consequently reclassified within the phylogenetic tree. In some cases, an SNP is found to be unreliable as a defining mutation and an SNP-based name is removed completely. For example, before 2005, R1b was synonymous with R-P25, which was later reclassified as R1b1; in 2016, R-P25 was removed completely as a defining SNP, due to a significant rate of back-mutation.[22]
This is the basic outline of R1b according to the ISOGG Tree as it stood on January 30, 2017.[1]
| 2017 ISOGG tree | |||||||||||||||||||||||||||||||||||||||||||||
|
R1b* (R-M343*)
R1b* – that is, males with M343, but no subsequent distinguishing SNP mutations – is extremely rare. The only population yet recorded with a definite significant proportion of R1b* are the Kurds of southeastern Kazakhstan with 13%.[6][23] However, more recently, a large study of Y-chromosome variation in Iran, revealed R1b* as high as 4.3% among Iranian sub-populations.[24]
While studies in 2005–08 suggested that R1b* may occur at high levels in Jordan, Egypt and Sudan, subsequent research indicates that the samples concerned most likely belong to the subclade R1b1a2 (R-V88), which is now concentrated among African populations, following back migration from Asia.[25][26][27][28][29] It remains a possibility that some or even most of these cases may be R1b* (R-M343*), R1a* (R-M420*), an otherwise undocumented branch of R1, and/or back-mutations of a marker, from a positive to a negative ancestral state,[30] constituting, in other words, undocumented subclades of R1b. Thus demonstrating the importance of testing for SNPs critical in identifying subclades.
A compilation of previous studies regarding the distribution of R1b can be found in Cruciani et al. (2010).[31] It is summarised in the table following. (It should be noted that Cruciani did not include some studies suggesting even higher frequencies of R1b1a1a2 [R-M269] in some parts of Western Europe.)
| Continent | Region | Sample size | Total R1b | R-P25 (unreliable marker for R1b1*) |
R-V88 (R1b1a2) | R-M269 (R1b1a1a2) | R-M73 (R1b1a1a1) |
|---|---|---|---|---|---|---|---|
| Africa | Northern Africa | 691 | 5.9% | 0.0% | 5.2% | 0.7% | 0.0% |
| Africa | Central Sahel Region | 461 | 23.0% | 0.0% | 23.0% | 0.0% | 0.0% |
| Africa | Western Africa | 123 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Africa | Eastern Africa | 442 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Africa | Southern Africa | 105 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Europe | Western Europeans | 465 | 57.8% | 0.0% | 0.0% | 57.8% | 0.0% |
| Europe | North-west Europeans | 43 | 55.8% | 0.0% | 0.0% | 55.8% | 0.0% |
| Europe | Central Europeans | 77 | 42.9% | 0.0% | 0.0% | 42.9% | 0.0% |
| Europe | North Eastern Europeans | 74 | 1.4% | 0.0% | 0.0% | 1.4% | 0.0% |
| Europe | Russians | 60 | 6.7% | 0.0% | 0.0% | 6.7% | 0.0% |
| Europe | Eastern Europeans | 149 | 20.8% | 0.0% | 0.0% | 20.8% | 0.0% |
| Europe | South-east Europeans | 510 | 13.1% | 0.0% | 0.2% | 12.9% | 0.0% |
| Asia | West Asians | 328 | 5.8% | 0.0% | 0.3% | 5.5% | 0.0% |
| Asia | South Asians | 288 | 4.8% | 0.0% | 0.0% | 1.7% | 3.1% |
| Asia | South-east Asians | 10 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Asia | North-east Asians | 30 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| Asia | East Asians | 156 | 0.6% | 0.0% | 0.0% | 0.6% | 0.0% |
| TOTAL | 5326 |
R1b1 (R-L278)
R1b1* or R-L278* is rare among modern males. However, it has been found in two skeletons from prehistoric Europe: a male from the Mesolithic Samara culture (a pre-Yamna people who lived immediately north of the Caspian Sea) buried in about 5650–5555 BCE, and a male from the early Neolithic Cardial culture, buried in about 5178–5066 BCE at the Els Trocs site in the Pyrenees (modern Aragon, Spain).[32]
Some examples described in older articles, for example two found in Turkey,[21] are now thought to be mostly in the more recently discovered sub-clade R1b1a2 (R-V88). Most examples of R1b therefore fall into subclades R1b1a2 (R-V88) or R1b1a (R-P297). Cruciani et al. in the large 2010 study found 3 cases amongst 1173 Italians, 1 out of 328 West Asians and 1 out of 156 East Asians.[31] Varzari found 3 cases in the Ukraine, in a study of 322 people from the Dniester-Carpathian Mountains region, who were P25 positive, but M269 negative.[33] Cases from older studies are mainly from Africa, the Middle East or Mediterranean, and are discussed below as probable cases of R1b1a2 (R-V88).
R1b1a (R-L754)
R1b1a (R-L754) was carried by an individual known as Villabruna 1, who lived c. 15,000 years BP in north-west Italy, and belonged to the Epigravettian culture.
Living individuals positive for L761, an equivalent to L754, have been found at high frequencies among the Toubou population inhabiting Chad (34%).[34]
R1b1a1 (R-L388)
R-L388, also known as R1b1a1 (L388/PF6468, L389/PF6531) appears to be rare or extinct in its basal form. Its subclades are also relatively rare and found in various parts of South West Asia, the Mediterranean basin and continental Europe.
R1b1a1a (R-P297)
The SNP marker P297 was recognised in 2008 as ancestral to the significant subclades M73 and M269, combining them into one cluster.[7] This had been given the phylogenetic name R1b1a1a (and, previously, R1b1a).
A majority of Eurasian R1b falls within this subclade, representing a very large modern population. Although P297 itself has not yet been much tested for, the same population has been relatively well studied in terms of other markers. Therefore, the branching within this clade can be explained in relatively high detail below. The skeleton of a male from a Chalcolithic Yamna burial in the Middle-Volga-Samara area, dated to around 3305–2925 BC, was found to possibly contain R1b1a* being P297 positive but L51 negative.[32]
R1b1a1a1 (R-M73)
R-M73 is reportedly the dominant haplogroup among the Kumandin of the Altai Republic in Russia.[35]
While early research into R-M73 claimed that it was significantly represented among the Hazara of Afghanistan and the Bashkirs of the Ural Mountains, this has apparently been overturned. For example, supporting material from a 2010 study by Behar et al. suggested that Sengupta et al. (2006) had misidentified Hazara individuals, who instead belonged to R2 or Q.[36][37][38] Likewise, most Bashkir males have been found to belong to U-152 (R1b1a1a2a1a2b) and some, mostly from south-eastern Bashkortostan, belonged to Haplogroup Q-M25 (Q1a1b) rather than R1b.
R1b1a1a2 (R-M269)
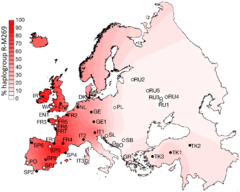
R-M269, or R1b1a1a2 (as of 2017) amongst other names,[39] is now the most common Y-DNA lineage in European males. It is carried by an estimated 110 million males in Europe.[9]
R-M269 has received significant scientific and popular interest due to its possible connection to the Indo-European expansion in Europe. Specifically the R-L23 (R-Z2103) subclade has been found to be prevalent in ancient DNA associated with the Yamna culture.[32] R-M269 likely originated in Western Asia and was present in Europe by the Neolithic period.[1][6][11][16] The distribution of subclades within Europe is substantially due to the various migrations of the Bronze and Iron Age. Western European populations are divided between the R-P312/S116 and R-U106/S21 subclades of R-M412 (R-L51). In Western Europe it is present but in generally much lower levels apart from "an instance of 27% in Switzerland's Upper Rhone Valley."[6]
Distribution of R-M269 in Europe increases in frequency from east to west. It peaks at the national level in Wales at a rate of 92%, at 82% in Ireland, 70% in Scotland, 68% in Spain, 60% in France (76% in Normandy), about 60% in Portugal, 53% in Italy,[6] 45% in Eastern England, 50% in Germany, 50% in the Netherlands, 42% in Iceland, and 43% in Denmark. R-M269 reaches levels as high as 95% in parts of Ireland. It has also been found at lower frequencies throughout central Eurasia,[40] but with relatively high frequency among the Bashkirs of the Perm region (84.0%).[41] This marker is present in China and India at frequencies of less than one percent. In Africa and adjoining islands, while R-V88 (R1b1a2) is more strongly represented, R-M269 appears to have been present at very high rates since prehistory. R-M269 has been found, for instance, at a rate of ~44% among remains dating from the 10th Century at Punta Azul, in the Canary Islands. These remains have been linked to the Bimbache (or Bimape), a subgroup of the Guanche.[42] In living males, it peaks in parts of North Africa, especially Algeria, at a rate of 10%.[43] In Sub-Saharan Africa, R-M269 appears to peak in Namibia, at a rate of 8% among Herero males.[29] (The table below lists in more detail the frequencies of M269 in regions in Asia, Europe, and Africa.)
Apart from undiverged, basal R-M269*, there are (as of 2017) two primary branches of R-M269:
- R-L23 (R1b1a1a2a; L23/PF6534/S141) and
- R-PF7558 (R1b1a1a2b; PF7558/PF7562.)
R-L23* and its subclades are by far the most common forms of R-M269. R-PF7558 is now rare, but has been reported anecdotally at low levels throughout Western Eurasia (Europe and the Middle East).[44]
R-L23 (Z2105/Z2103; a.k.a. R1b1a1a2a) has been reported among the peoples of the Idel-Ural (by Trofimova et al. 2015): 21 out of 58 (36.2%) of Burzyansky District Bashkirs, 11 out of 52 (21.2%) of Udmurts, 4 out of 50 (8%) of Komi, 4 out of 59 (6.8%) of Mordvins, 2 out of 53 (3.8%) of Besermyan and 1 out of 43 (2.3%) of Chuvash were R1b-L23.[45]
Subclades within the paragroup R-M269(xL23) – that is, R-M269* and/or R-PF7558 – appear to be found at their highest frequency in the central Balkans, especially Kosovo with 7.9%, Macedonia 5.1% and Serbia 4.4%.[6] Unlike most other areas with significant percentages of R-L23, Kosovo, Poland and the Bashkirs of south-east Bashkortostan are notable in having a high percentage of R-L23(xM412) also known as R1b1a1a2a(xR1b1a1a2a1) – at rates of 11.4% (Kosovo), 2.4% (Poland) and 2.4% south-east Bashkortostan.[6] (This Bashkir population is also notable for its high level of R-M73 (R1b1a1a1), at 23.4%.[6]) Five individuals out of 110 tested in the Ararat Valley of Armenia belonged to R-M269(xL23) and 36 to R-L23*, with none belonging to known subclades of L23.[46]
In 2009, DNA extracted from the femur bones of 6 skeletons in an early-medieval burial place in Ergolding (Bavaria, Germany) dated to around AD 670 yielded the following results: 4 were found to be haplogroup R1b with the closest matches in modern populations of Germany, Ireland and the USA while 2 were in Haplogroup G2a.[47]
The following gives a summary of most of the studies which specifically tested for M269, showing its distribution (as a percentage of total population) in Europe, North Africa, the Middle East and Central Asia as far as China and Nepal.
The phylogeny of R-M269 according to ISOGG 2017:
| M269/PF6517 |
| |||||||||||||||||||||||||||||||||||||||||||||
R1b1a1b (R-V1636)
R-V1636 (R1b1a1b) has no known subclades, is rare and has been found mostly in the broader Mediterranean region.
R1b1a2 (R-V88)
R1b1a2 (PF6279/V88; previously R1b1c) is defined by the presence of SNP marker V88, the discovery of which was announced in 2010 by Cruciani et al.[31] Apart from individuals in southern Europe and Western Asia, the majority of R-V88 was found in the Sahel among populations speaking Afroasiatic languages of the Chadic branch:
| Region | Population | Country | Language | N | Total% | R1b1c (R-V88) | R1b1a1a2 (R-M269) | R1b1c* (R-V88*) | R1b1c3 (R-V69) |
|---|---|---|---|---|---|---|---|---|---|
| N Africa | Composite | Morocco | AA | 338 | 0.0% | 0.3% | 0.6% | 0.3% | 0.0% |
| N Africa | Mozabite Berbers | Algeria | AA/Berber | 67 | 3.0% | 3.0% | 0.0% | 3.0% | 0.0% |
| N Africa | Northern Egyptians | Egypt | AA/Semitic | 49 | 6.1% | 4.1% | 2.0% | 4.1% | 0.0% |
| N Africa | Berbers from Siwa | Egypt | AA/Berber | 93 | 28.0% | 26.9% | 1.1% | 23.7% | 3.2% |
| N Africa | Baharia | Egypt | AA/Semitic | 41 | 7.3% | 4.9% | 2.4% | 0.0% | 4.9% |
| N Africa | Gurna Oasis | Egypt | AA/Semitic | 34 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| N Africa | Southern Egyptians | Egypt | AA/Semitic | 69 | 5.8% | 5.8% | 0.0% | 2.9% | 2.9% |
| C Africa | Songhai | Niger | NS/Songhai | 10 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| C Africa | Fulbe | Niger | NC/Atlantic | 7 | 14.3% | 14.3% | 0.0% | 14.3% | 0.0% |
| C Africa | Tuareg | Niger | AA/Berber | 22 | 4.5% | 4.5% | 0.0% | 4.5% | 0.0% |
| C Africa | Ngambai | Chad | NS/Sudanic | 11 | 9.1% | 9.1% | 0.0% | 9.1% | 0.0% |
| C Africa | Hausa | Nigeria (North) | AA/Chadic | 10 | 20.0% | 20.0% | 0.0% | 20.0% | 0.0% |
| C Africa | Fulbe | Nigeria (North) | NC/Atlantic | 32 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| C Africa | Yoruba | Nigeria (South) | NC/Defoid | 21 | 4.8% | 4.8% | 0.0% | 4.8% | 0.0% |
| C Africa | Ouldeme | Cameroon (Nth) | AA/Chadic | 22 | 95.5% | 95.5% | 0.0% | 95.5% | 0.0% |
| C Africa | Mada | Cameroon (Nth) | AA/Chadic | 17 | 82.4% | 82.4% | 0.0% | 76.5% | 5.9% |
| C Africa | Mafa | Cameroon (Nth) | AA/Chadic | 8 | 87.5% | 87.5% | 0.0% | 25.0% | 62.5% |
| C Africa | Guiziga | Cameroon (Nth) | AA/Chadic | 9 | 77.8% | 77.8% | 0.0% | 22.2% | 55.6% |
| C Africa | Daba | Cameroon (Nth) | AA/Chadic | 19 | 42.1% | 42.1% | 0.0% | 36.8% | 5.3% |
| C Africa | Guidar | Cameroon (Nth) | AA/Chadic | 9 | 66.7% | 66.7% | 0.0% | 22.2% | 44.4% |
| C Africa | Massa | Cameroon (Nth) | AA/Chadic | 7 | 28.6% | 28.6% | 0.0% | 14.3% | 14.3% |
| C Africa | Other Chadic | Cameroon (Nth) | AA/Chadic | 4 | 75.0% | 75.0% | 0.0% | 25.0% | 50.0% |
| C Africa | Shuwa Arabs | Cameroon (Nth) | AA/Semitic | 5 | 40.0% | 40.0% | 0.0% | 40.0% | 0.0% |
| C Africa | Kanuri | Cameroon (Nth) | NS/Saharan | 7 | 14.3% | 14.3% | 0.0% | 14.3% | 0.0% |
| C Africa | Fulbe | Cameroon (Nth) | NC/Atlantic | 18 | 11.1% | 11.1% | 0.0% | 5.6% | 5.6% |
| C Africa | Moundang | Cameroon (Nth) | NC/Adamawa | 21 | 66.7% | 66.7% | 0.0% | 14.3% | 52.4% |
| C Africa | Fali | Cameroon (Nth) | NC/Adamawa | 48 | 20.8% | 20.8% | 0.0% | 10.4% | 10.4% |
| C Africa | Tali | Cameroon (Nth) | NC/Adamawa | 22 | 9.1% | 9.1% | 0.0% | 4.5% | 4.5% |
| C Africa | Mboum | Cameroon (Nth) | NC/Adamawa | 9 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| C Africa | Composite | Cameroon (Sth) | NC/Bantu | 90 | 0.0% | 1.1% | 0.0% | 1.1% | 0.0% |
| C Africa | Biaka Pygmies | CAR | NC/Bantu | 33 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% |
| W Africa | Composite | — | 123 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | |
| E Africa | Composite | — | 442 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | |
| S Africa | Composite | — | 105 | 0.0% | 0.0% | 0.0% | 0.0% | 0.0% | |
| TOTAL | 1822 |
| V88 |
| ||||||||||||
As can be seen in the above data table, R1b1c is found in northern Cameroon in west central Africa at a very high frequency, where it is considered to be caused by a pre-Islamic movement of people from Eurasia.[29][48]
R1b1a2a (R-M18)
R1b1a2a is a sub-clade of R-V88, which is defined by the presence of SNP marker M18.[7] It has been found only at low frequencies in samples from Sardinia[40][49] and Lebanon.[50]
R1b1b (R-PH155)
The other secondary branch of R1b is R-PH155 (R1b1b), which is extremely rare and defined by the presence of PH155.[51] ISOGG (2017) cites two primary branches: R-M335 (R1b1b1) and R-PH200 (R1b1b2).
Another defining SNP of R1b1b, M335, was first documented in 2004, when an example was discovered in Turkey, which was classified at that time as R1b4.[21]
Other examples of R-PH155 have been found in Bahrain, Bhutan, Tajikistan and Xinjiang.
In popular culture
- Bryan Sykes, in his 2006 book Blood of the Isles, gives the members – and the notional founding patriarch – of R1b the name "Oisín".
- Stephen Oppenheimer, in his 2007 book Origins of the British, gives the R1b patriarch the Basque name "Ruisko" in honour of what Oppenheimer believed to be the Iberian origin of R1b.
- A filmmaker named Artem Lukichev created (circa 2009), a 14-minute animated film based on a Bashkir epic from the Ural Mountains, relating the epic to the emergence and geographical expansion of R1a and R1b.[52]
- DNA tests that assisted in the identification of Czar Nicholas II of Russia found that he belonged to R1b.[53] This may suggest that the later Czars of the House of Romanov , descended in the male line from the House of Holstein-Gottorp (which originated in Schleswig-Holstein) are also members of R1b.
See also
| Wikimedia Commons has media related to Haplogroup R1b of Y-DNA. |
- Human Y-chromosome DNA haplogroup
- Atlantic Modal Haplotype
- Genealogical DNA test
- Prehistoric Europe
- Y-DNA haplogroups in populations of Europe
| Phylogenetic tree of human Y-chromosome DNA haplogroups [χ 1][χ 2] | |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| "Y-chromosomal Adam" | |||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0-T [χ 3] | ||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1 [χ 4] | ||||||||||||||||||||||||||||||||||||||||||||||||
| A1a | A1b | ||||||||||||||||||||||||||||||||||||||||||||||||
| A1b1 | BT | ||||||||||||||||||||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | ||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | ||||||||||||||||||||||||||||||||||||||||||||||
| F1 | F2 | F3 | GHIJK | ||||||||||||||||||||||||||||||||||||||||||||||
| G | HIJK | ||||||||||||||||||||||||||||||||||||||||||||||||
| IJK | H | ||||||||||||||||||||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||||||||||||||||||||
| I | J | LT [χ 5] | K2 | ||||||||||||||||||||||||||||||||||||||||||||||
| L | T [χ 6] | K2a [χ 7] | K2b [χ 8] | K2c | K2d | K2e [χ 9] | |||||||||||||||||||||||||||||||||||||||||||
| K2a1 | K2b1 [χ 10] | P [χ 11] | |||||||||||||||||||||||||||||||||||||||||||||||
| NO | S [χ 12] | M [χ 13] | P1 | P2 | |||||||||||||||||||||||||||||||||||||||||||||
| NO1 | Q | R | |||||||||||||||||||||||||||||||||||||||||||||||
| N | O | ||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
References
- 1 2 3 International Society of Genetic Genealogy (ISOGG) – Y-DNA Haplogroup R and its Subclades
- 1 2 Karafet, Tatiana; Mendez, Fernando; Sudoyo, Herawati (2014). "Improved phylogenetic resolution and rapid diversification of Y-chromosome haplogroup K-M526 in Southeast Asia". Nature. 23: 369–373. PMC 4326703
 . PMID 24896152. doi:10.1038/ejhg.2014.106.
. PMID 24896152. doi:10.1038/ejhg.2014.106. - ↑ Massive migration from the steppe is a source for Indo-European languages in Europe, Haak et al, 2015
- ↑ Population genomics of Bronze Age Eurasia, Allentoft et al, 2015
- ↑ Eight thousand years of natural selection in Europe, Mathieson et al, 2015
- 1 2 3 4 5 6 7 8 9 10 11 12 Myres, Natalie; Rootsi, Siiri; Lin, Alice A; Järve, Mari; King, Roy J; Kutuev, Ildus; Cabrera, Vicente M; Khusnutdinova, Elza K; et al. (2010). "A major Y-chromosome haplogroup R1b Holocene effect in Central and Western Europe". European Journal of Human Genetics. 19 (1): 95–101. PMC 3039512 . PMID 20736979. doi:10.1038/ejhg.2010.146
- 1 2 3 Karafet, TM; Mendez, FL; Meilerman, MB; Underhill, PA; Zegura, SL; Hammer, MF (2008). "New binary polymorphisms reshape and increase resolution of the human Y chromosomal haplogroup tree". Genome Research. 18 (5): 830–8. PMC 2336805 . PMID 18385274. doi:10.1101/gr.7172008.
- ↑ Semino O, Passarino G, Oefner PJ, Lin AA, Arbuzova S, Beckman LE, De Benedictis G, Francalacci P, Kouvatsi A, et al. (2000). "The genetic legacy of paleolithic Homo sapiens sapiens in extant Europeans: a Y chromosome perspective". Science. 290 (5494): 1155–59. PMID 11073453. doi:10.1126/science.290.5494.1155.
- 1 2 3 4 Balaresque, Patricia; Bowden, Georgina R.; Adams, Susan M.; Leung, Ho-Yee; King, Turi E.; et al. (2010). Penny, David, ed. "A Predominantly Neolithic Origin for European Paternal Lineages". PLOS Biology. Public Library of Science. 8 (1): e1000285. PMC 2799514 . PMID 20087410. doi:10.1371/journal.pbio.1000285. Retrieved August 19, 2014.
- ↑ Sjödin, Per; François, Olivier (2011). Lalueza-Fox, Carles, ed. "Wave-of-Advance Models of the Diffusion of the Y Chromosome Haplogroup R1b1b2 in Europe". PLoS ONE. 6 (6:e21592): e21592. PMC 3123369 . PMID 21720564. doi:10.1371/journal.pone.0021592
- 1 2 3 B. Arredi; E. S. Poloni; C. Tyler-Smith (2007). "The peopling of Europe". In Crawford, Michael H. Anthropological genetics: theory, methods and applications. Cambridge, UK: Cambridge University Press. p. 394. ISBN 0-521-54697-4.
- ↑ Morelli, Laura; Contu, Daniela; Santoni, Federico; Whalen, Michael B.; Francalacci, Paolo; Cucca, Francesco (2010). Lalueza-Fox, Carles, ed. "A Comparison of Y-Chromosome Variation in Sardinia and Anatolia Is More Consistent with Cultural Rather than Demic Diffusion of Agriculture". PLoS ONE. 5 (4): e10419. PMC 2861676 . PMID 20454687. doi:10.1371/journal.pone.0010419
- ↑ http://www.pnas.org/content/95/15/9053.full.pdf+html
- ↑ Maglio, Michael R. (2014-08-14). "Biogeographical Evidence for the Iberian Origins of R1b-L278 via Haplotype Aggregation (2014)". Origins DNA. Retrieved 2014-11-12.
- ↑ Chiaroni, J; Underhill, P; Cavalli-Sforza, L.L. (2009). "Y chromosome diversity, human expansion, drift and cultural evolution". PNAS. 106 (48): 20174:20179. PMC 2787129 . PMID 19920170. doi:10.1073/pnas.0910803106
- 1 2 Cruciani; Trombetta, Beniamino; Antonelli, Cheyenne; Pascone, Roberto; Valesini, Guido; Scalzi, Valentina; Vona, Giuseppe; Melegh, Bela; et al. (2010). "Strong intra- and inter-continental differentiation revealed by Y chromosome SNPs M269, U106 and U152". Forensic Science International: Genetics. 5 (3): e49. PMID 20732840. doi:10.1016/j.fsigen.2010.07.006
- ↑ Lee, Esther J.; et al. (3 May 2012). "Emerging genetic patterns of the european neolithic: Perspectives from a late neolithic bell beaker burial site in Germany". American Journal of Physical Anthropology. 148 (4): 571–9. PMID 22552938. doi:10.1002/ajpa.22074.
- ↑ Lacan, Marie; et al. (May 2, 2011). "Ancient DNA reveals male diffusion through the Neolithic Mediterranean route". Proceedings of the National Academy of Sciences of the United States of America. 108 (24): 9788–91. PMC 3116412 . PMID 21628562. doi:10.1073/pnas.1100723108.
- ↑ Haak, Wolfgang; et al. (November 2010). Penny, David, ed. "Ancient DNA from European Early Neolithic Farmers Reveals Their Near Eastern Affinities". PLoS Biology. 8 (11): e1000536. PMC 2976717 . PMID 21085689. doi:10.1371/journal.pbio.1000536.
- 1 2 eupedia.com/genetics
- 1 2 3 Cinnioğlu, C; King, R; Kivisild, T; Kalfoğlu, E; Atasoy, S; Cavalleri, GL; Lillie, AS; Roseman, CC; et al. (2004). "Excavating Y-chromosome haplotype strata in Anatolia" (PDF). Human Genetics. 114 (2): 127–48. PMID 14586639. doi:10.1007/s00439-003-1031-4.
- ↑ ISOGG, Y-DNA Haplogroup R and its Subclades - 2016 (8 February 2017).
- ↑ Supplementary Table S4 (xls 62K) -> http://www.nature.com/ejhg/journal/v19/n1/suppinfo/ejhg2010146s1.html
- ↑ PLOS ONE: Ancient Migratory Events in the Middle East: New Clues from the Y-Chromosome Variation of Modern Iranians
- ↑ Flores et. al. (2005) found that 20 out of all 146 men tested (13.7%) – including 20 out of 45 men tested from the Dead Sea area of Jordan – were positive for M173 (R1), and negative for both the R1a markers SRY10831.2 and M17, as well as the R1b subclades P25 (R1b1) and M269 (R1b1a2), a study indicates that they are R1b2 (R-V88). Wood et al. (2005) reported two Egyptian cases of R1-M173 that were negative for SRY10831 (R1a1) and P25 (R1b1), out of a sample of 1,122 males from African countries, including 92 from Egypt. Hassan et al. (2008) found an equally surprising 14 out of 26 (54%) of Sudanese Fula people who were M173+ and P25-
- ↑ Natalie M Myres, Siiri Rootsi, Alice A Lin, Mari Järve, Roy J King, Ildus Kutuev, Vicente M Cabrera, Elza K Khusnutdinova, Andrey Pshenichnov, Bayazit Yunusbayev, Oleg Balanovsky, Elena Balanovska, Pavao Rudan, Marian Baldovic, Rene J Herrera, Jacques Chiaroni, Julie Di Cristofaro, Richard Villems, Toomas Kivisild & Peter A Underhill, 2010, "A major Y-chromosome haplogroup R1b Holocene era founder effect in Central and Western Europe", Nature 2010, v. 19, no. 1 (28 May 2016)
- ↑ Flores, C; Maca-Meyer, N; Larruga, JM; Cabrera, VM; Karadsheh, N; Gonzalez, AM (2005). "Isolates in a corridor of migrations: a high-resolution analysis of Y-chromosome variation in Jordan". Journal of Human Genetics. 50 (9): 435–41. PMID 16142507. doi:10.1007/s10038-005-0274-4.
- ↑ Hassan, HY; Underhill, PA; Cavalli-Sforza, LL; Ibrahim, ME (2008). "Y-chromosome variation among Sudanese: restricted gene flow, concordance with language, geography, and history" (PDF). American Journal of Physical Anthropology. 137 (3): 316–23. PMID 18618658. doi:10.1002/ajpa.20876.
13/32
- 1 2 3 Wood, ET; Stover, DA; Ehret, C; Destro-Bisol, G; Spedini, G; Mcleod, H; Louie, L; Bamshad, M; et al. (2005). "Contrasting patterns of Y chromosome and mtDNA variation in Africa: evidence for sex-biased demographic processes" (PDF). European Journal of Human Genetics. 13 (7): 867–76. PMID 15856073. doi:10.1038/sj.ejhg.5201408. Archived from the original (PDF) on June 26, 2008.
- ↑ Adams, SM; King, TE; Bosch, E; Jobling, MA (2006). "The case of the unreliable SNP: recurrent back-mutation of Y-chromosomal marker P25 through gene conversion". Forensic Science International. 159 (1): 14–20. PMID 16026953. doi:10.1016/j.forsciint.2005.06.003.
- 1 2 3 Cruciani; Trombetta, B; Sellitto, D; Massaia, A; Destro-Bisol, G; Watson, E; Beraud Colomb, E; Dugoujon, JM; et al. (2010). "Human Y chromosome haplogroup R-V88: a paternal genetic record of early mid Holocene trans-Saharan connections and the spread of Chadic languages". European Journal of Human Genetics. 18 (7): 800–7. PMC 2987365 . PMID 20051990. doi:10.1038/ejhg.2009.231.
- 1 2 3 Haak, Wolfgang; Lazaridis, Iosif (February 10, 2015). "Massive migration from the steppe is a source for Indo-European languages in Europe". bioRxiv 013433 .
- ↑ Varzari, Alexander (2006). "Population History of the Dniester-Carpathians: Evidence from Alu Insertion and Y-Chromosome Polymorphisms" (PDF). Dissertation der Fakultät für Biologie der Ludwig-Maximilians-Universität München.
- ↑ Haber, Marc; et al. (2016). "Chad Genetic Diversity Reveals an African History Marked by Multiple Holocene Eurasian Migrations". American Journal of Human Genetics. 99 (6): 1316–1324. Retrieved 27 June 2017. - Y-chromosomal haplogroup frequencies on Table S.4
- ↑ Dulik, MC; Zhadanov, SI; Osipova, LP; Askapuli, A; Gau, L; Gokcumen, O; Rubinstein, S; Schurr, TG (2012). "Mitochondrial DNA and Y chromosome variation provides evidence for a recent common ancestry between Native Americans and Indigenous Altaians". Am. J. Hum. Genet. 90: 229–46. PMC 3276666 . PMID 22281367. doi:10.1016/j.ajhg.2011.12.014.
- ↑ http://www.nature.com/nature/journal/v466/n7303/extref/nature09103-s1.pdf
- ↑ European Journal of Human Genetics - Supplementary Information for article: A major Y-chromosome haplogroup R1b Holocene era founder effect in Central and Western Europe
- ↑ Sengupta, S; Zhivotovsky, LA; King, R; et al. (February 2006). "Polarity and temporality of high-resolution y-chromosome distributions in India identify both indigenous and exogenous expansions and reveal minor genetic influence of Central Asian pastoralists". Am. J. Hum. Genet. 78: 202–21. PMC 1380230 . PMID 16400607. doi:10.1086/499411.
- ↑ R1b1a1a2 (R-M269) was previously R1b1a2, From 2003 to 2005, what is now R1b1a2 was designated R1b3. From 2005 to 2008, it was R1b1c. From 2008 to 2011, it was R1b1b2.
- 1 2 Peter A. Underhill, Peidong Shen, Alice A. Lin et al., "Y chromosome sequence variation and the history of human populations", Nature Genetics, Volume 26, November 2000
- ↑ A. S. Lobov et al. (2009), "Structure of the Gene Pool of Bashkir Subpopulations" (original text in Russian)
- ↑ Ordóñez, A. C., Fregel, R., Trujillo-Mederos, A., Hervella, M., de-la-Rúa, C., & Arnay-de-la-Rosa, M. (2017). "Genetic studies on the prehispanic population buried in Punta Azul cave (El Hierro, Canary Islands)". Journal of Archaeological Science. 78: 20–28. doi:10.1016/j.jas.2016.11.004. Retrieved 16 February 2017.
- ↑ Analysis of Y-chromosomal SNP haplogroups and STR haplotypes in an Algerian population sample
- ↑ https://www.familytreedna.com/public/R1bBasalSubclades
- ↑ Трофимова Натал'я Вадимовна (Feb. 2015), "Изменчивость Митохондриальной ДНК и Y-Хромосомы в Популяциях Волго-Уральского Региона" ("Mitochondrial DNA variation and the Y-chromosome in the population of the Volga-Ural Region"). Автореферат. диссертации на соискание ученой степени кандидата биологических наук. Уфа – 2015.
- ↑ Kristian, J Herrera; Lowery, Robert K; Hadden, Laura. "Haplotype diversity, variance and time estimations for Haplogroup R1b". European Journal of Human Genetics. 20 (3): Table 3. PMC 3286660 . PMID 22085901. doi:10.1038/ejhg.2011.192.
- ↑ Vanek, Daniel; Saskovat and Koch (June 2009). "Kinship and Y-Chromosome Analysis of 7th Century Human Remains: Novel DNA Extraction and Typing Procedure for Ancient Material". Croatian Medical Journal. 3. 50 (3): 286–295. PMC 2702742 . PMID 19480023. doi:10.3325/cmj.2009.50.286.
- ↑ Cruciani, F; Santolamazza, P; Shen, P; Macaulay, V; Moral, P; Olckers, A; Modiano, D; Holmes, S; et al. (2002). "A Back Migration from Asia to Sub-Saharan Africa Is Supported by High-Resolution Analysis of Human Y-Chromosome Haplotypes". American Journal of Human Genetics. 70 (5): 1197–214. PMC 447595 . PMID 11910562. doi:10.1086/340257., pp. 13–14
- ↑ Contu, D; Morelli; Santoni; Foster; Francalacci; Cucca (2008). "Y-Chromosome Based Evidence for Pre-Neolithic Origin of the Genetically Homogeneous but Diverse Sardinian Population: Inference for Association Scans". PLoS ONE. 3 (1): e1430. PMC 2174525 . PMID 18183308. doi:10.1371/journal.pone.0001430.
- ↑ Zalloua, PA; Xue, Y; Khalife, J; Makhoul, N; Debiane, L; Platt, DE; Royyuru, AK; Herrera, RJ; Hernanz, DF; et al. (2008). "Y-Chromosomal Diversity in Lebanon Is Structured by Recent Historical Events". American Journal of Human Genetics. 82 (4): 873–82. PMC 2427286 . PMID 18374297. doi:10.1016/j.ajhg.2008.01.020.
- ↑ ISOGG, Y-DNA Haplogroup R and its Subclades - 2017 (1 August 2017).
- ↑ About R1a and R1b from Ural epic story. Artem Lukichev (c)
- ↑ Coble MD, Loreille OM, Wadhams MJ, Edson SM, Maynard K, et al. (2009). "Mystery Solved: The Identification of the Two Missing Romanov Children Using DNA Analysis". PLoS ONE. 4 (3): e4838. PMC 2652717 . PMID 19277206. doi:10.1371/journal.pone.0004838.
Unannotated references
- Nature Article - 日本語要約 - The genetic history of Ice Age Europe - Qiaomei Fu, Cosimo Posth, Mateja Hajdinjak, Martin Petr, Swapan Mallick, Daniel Fernandes, Anja Furtwängler, Wolfgang Haak, Matthias Meyer, Alissa Mittnik, Birgit Nickel, Alexander Peltzer, Nadin Rohland, Viviane Slon, Sahra Talamo, Iosif Lazaridis, Mark Lipson, Iain Mathieson, Stephan Schiffels, Pontus Skoglund, Anatoly P. Derevianko, Nikolai Drozdov, Vyacheslav Slavinsky, Alexander Tsybankov, Renata Grifoni Cremonesi, Francesco Mallegni, Bernard Gély, Eligio Vacca, Manuel R. González Morales, Lawrence G. Straus, Christine Neugebauer-Maresch, Maria Teschler-Nicola, Silviu Constantin, Oana Teodora Moldovan, Stefano Benazzi, Marco Peresani, Donato Coppola, Martina Lari, Stefano Ricci, Annamaria Ronchitelli, Frédérique Valentin, Corinne Thevenet, Kurt Wehrberger, Dan Grigorescu, Hélène Rougier, Isabelle Crevecoeur, Damien Flas, Patrick Semal, Marcello A. Mannino, Christophe Cupillard, Hervé Bocherens, Nicholas J. Conard, Katerina Harvati, Vyacheslav Moiseyev, Dorothée G. Drucker, Jiří Svoboda, Michael P. Richards, David Caramelli, Ron Pinhasi, Janet Kelso, Nick Patterson, Johannes Krause, Svante Pääbo & David Reich - Nature 534, 200–205 (09 June 2016) doi:10.1038/nature17993 Received 18 December 2015, Accepted 12 April 2016, Published online 02 May 2016 (in particular RE: Epigravettian R1b find)