Protein disulfide-isomerase
| Protein disulfide-isomerase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| EC number | 5.3.4.1 | ||||||||
| CAS number | 37318-49-3 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / EGO | ||||||||
| |||||||||
| protein disulfide isomerase family A, member 2 | |
|---|---|
| Identifiers | |
| Symbol | PDIA2 |
| Alt. symbols | PDIP |
| Entrez | 64714 |
| HUGO | 14180 |
| OMIM | 608012 |
| RefSeq | NM_006849 |
| UniProt | Q13087 |
| Other data | |
| Locus | Chr. 16 p13.3 |
| protein disulfide isomerase family A, member 3 | |
|---|---|
| Identifiers | |
| Symbol | PDIA3 |
| Alt. symbols | GRP58 |
| Entrez | 2923 |
| HUGO | 4606 |
| OMIM | 602046 |
| RefSeq | NM_005313 |
| UniProt | P30101 |
| Other data | |
| Locus | Chr. 15 q15 |
| protein disulfide isomerase family A, member 4 | |
|---|---|
| Identifiers | |
| Symbol | PDIA4 |
| Entrez | 9601 |
| HUGO | 30167 |
| RefSeq | NM_004911 |
| UniProt | P13667 |
| Other data | |
| Locus | Chr. 7 q35 |
| protein disulfide isomerase family A, member 5 | |
|---|---|
| Identifiers | |
| Symbol | PDIA5 |
| Entrez | 10954 |
| HUGO | 24811 |
| RefSeq | NM_006810 |
| UniProt | Q14554 |
| Other data | |
| EC number | 5.3.4.1 |
| Locus | Chr. 3 q21.1 |
| protein disulfide isomerase family A, member 6 | |
|---|---|
| Identifiers | |
| Symbol | PDIA6 |
| Alt. symbols | TXNDC7 |
| Entrez | 10130 |
| HUGO | 30168 |
| RefSeq | NM_005742 |
| UniProt | Q15084 |
| Other data | |
| Locus | Chr. 2 p25.1 |
Protein disulfide isomerase, or PDI, is an enzyme in the endoplasmic reticulum (ER) in eukaryotes and the periplasm of bacteria that catalyzes the formation and breakage of disulfide bonds between cysteine residues within proteins as they fold.[1][2][3] This allows proteins to quickly find the correct arrangement of disulfide bonds in their fully folded state, and therefore the enzyme acts to catalyze protein folding.
Structure
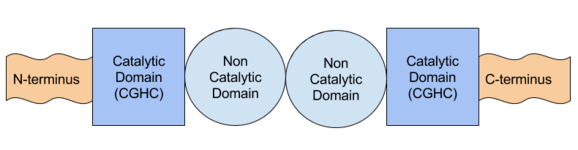
Protein disulfide-isomerase has two catalytic thioredoxin-like domains (active sites), each containing the canonical CGHC motif, and two non catalytic domains.[4][5][6] This structure is similar to the structure of enzymes responsible for oxidative folding in the intermembrane space of the mitochondria; an example of this is mitochondrial IMS import and assembly (Mia40), which has 2 catalytic domains that contain a CX9C, which is similar to the CGHC domain of PDI.[7]
Function
Protein folding
PDI displays oxidoreductase and isomerase properties, both of which depend on the type of substrate that binds to protein disulfide-isomerase and changes in protein disulfide-isomerase's redox state.[4] These types of activities allow for oxidative folding of proteins. Oxidative folding involves the oxidation of reduced cysteine residues of nascent proteins; upon oxidation of these cysteine residues, disulfide bridges are formed, which stabilizes proteins and allows for native structures (namely tertiary and quaternary structures).[5]
Regular Oxidative Folding Mechanism and Pathway
PDI is specifically responsible for folding proteins for the ER, which are first translated in the cytosol by free ribosomes.[6] For each unfolded protein, the first cysteine residue binds to either active site (CGHC motif) of protein disulfide-isomerase to form an enzyme substate complex; a second cysteine residue binds to the complex, whereby after subsequent catalysis, a stable disulfide bridge on the substrate is formed, leaving protein disulfide-isomerase's cysteine residues reduced.[4][5]
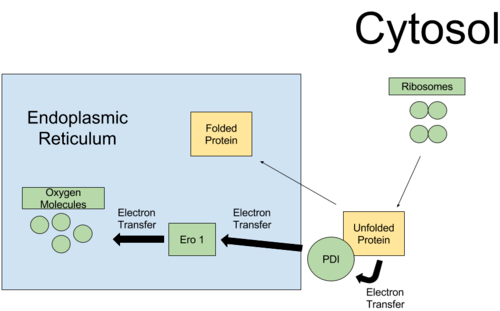
Afterwards, PDI can be regenerated to its oxidized form in the endoplasmic reticulum by transferring electrons to reoxidizing proteins such ER Oxidoreductin 1 (Ero 1), VKOR (vitamin K epoxide reductase), glutathione peroxidase (Gpx7/8), and PrxIV (peroxiredoxin IV).[4][8][9][6] Ero1 is thought to be the main reoxidizing protein of PDI, and the pathway of reoxidation of PDI for Ero1 is more understood than that of other proteins.[9] Ero1 accepts electrons from PDI and donates these electrons to oxygen molecules in the ER, which leads to the formation of hydrogen peroxide.[9]
Misfolded Protein Mechanism
The reduced (dithiol) form of protein disulfide-isomerase is able to catalyze a reduction of misformed disulfide bridge of a particular substrate through either reductase activity or isomerase activity.[10] For the reductase method, misfolded substrate disulfide bonds are converted to reduced cysteine residues by the transfer of electrons from glutathione and NADPH. Afterwards, normal oxidative folding occurs to the correct pairs of substrate cysteine residues, leading to properly folded proteins. For the isomerase method, intramolecular rearrangement of substrate functional groups is catalyzed near the N terminus of each active site.[4] Therefore, protein disulfide-isomerase is capable of catalyzing the post-translational modification disulfide exchange.
Redox signaling
In the chloroplasts of the unicellular algae Chlamydomonas reinhardtii the protein disulfide-isomerase RB60 serves as a redox sensor component of an mRNA-binding protein complex implicated in the photoregulation of the translation of psbA, the RNA encoding for the photosystem II core protein D1. Protein disulfide-isomerase has also been suggested to play a role in the formation of regulatory disulfide bonds in chloroplasts.[11]
Other functions
Immune System
Protein disulfide-isomerase helps load antigenic peptides into MHC class I molecules. These molecules (MHC I) are related to the peptide presentation by antigen-presenting cells in the immune response.
Protein disulfide-isomerase has been found to be involved in the breaking of bonds on the HIV gp120 protein during HIV infection of CD4 positive cells, and is required for HIV infection of lymphocytes and monocytes.[12] Some studies have shown it to be available for HIV infection on the surface of the cell clustered around the CD4 protein. Yet conflicting studies have shown that it is not available on the cell surface, but instead is found in significant amounts in the blood plasma.
Chaperone Activity
Another major function of protein disulfide-isomerase relates to its activity as a chaperone; its b' domain aids in the binding of misfolded protein for subsequent degradation.[4] This is regulated by three ER membrane proteins, Protein Kinase RNA-like endoplasmic reticulum kinase (PERK), inositol-requiring kinase 1 (IRE1), and activating transcription factor 6 (ATF6).[4][13] They respond to high levels of misfolded proteins in the ER through intracellular signaling cascades that can activate PDI's chaperone activity.[4] These signals can also inactivate translation of these misfolded proteins, because the cascade travels from the ER to the nucleus.[4]
Activity assays
Insulin Turbidity Assay: protein disulfide-isomerase breaks the two disulfide bonds between two insulin (a and b) chains that results in precipitation of b chain. This precipitation can be monitored at 650 nm, which is indirectly used to monitor protein disulfide-isomerase activity.[14] Sensitivity of this assay is in micromolar range.
ScRNase assay: protein disulfide-isomerase converts scrambled (inactive) RNase into native (active) RNase that further acts on its substrate.[15] The sensitivity is in micromolar range.
Di-E-GSSG assay: This is the fluorometric assay that can detect picomolar quantities of protein disulfide-isomerase and therefore is the most sensitive assay to date for detecting protein disulfide-isomerase activity.[16] Di-E-GSSG has two eosin molecules attached to oxidized glutathione (GSSG). The proximity of eosin molecules leads to the quenching of its fluorescence. However, upon breakage of disulfide bond by protein disulfide-isomerase, fluorescence increases 70-fold.
Stress and Inhibition
Effects of Nitrosative Stress
Redox dysregulation leads to increases in nitrosative stress in the endoplasmic reticulum. Such adverse changes in the normal cellular environment of susceptible cells, such as neurons, leads to nonfunctioning thiol-containing enzymes.[13] More specifically, protein disulfide-isomerase can no longer fix misfolded proteins once its thiol group in its active site has a nitric monoxide group attached to it; as a result, accumulation of misfolded proteins occurs in neurons, which has been associated with the defelopment of neurodegenerative diseases such as Alzheimer's Disease and Parkinson's Disease.[4][13]
Inhibition
Due to the role of protein disulfide-isomerase in a number of disease states, small molecule inhibitors of protein disulfide-isomerase have been developed. These molecules can either target the active site of protein disulfide-isomerase irreversibly [17] or reversibly.[18]
It has been shown that protein disulfide-isomerase activity is inhibited by red wine and grape juice, which could be the explanation for the French Paradox.[19]
Members
Human genes encoding protein disulfide isomerases include:[3][20][21]
References
- ↑ Wilkinson B, Gilbert HF (Jun 2004). "Protein disulfide isomerase". Biochimica et Biophysica Acta. 1699 (1–2): 35–44. PMID 15158710. doi:10.1016/j.bbapap.2004.02.017.
- ↑ Gruber CW, Cemazar M, Heras B, Martin JL, Craik DJ (Aug 2006). "Protein disulfide isomerase: the structure of oxidative folding". Trends in Biochemical Sciences. 31 (8): 455–64. PMID 16815710. doi:10.1016/j.tibs.2006.06.001.
- 1 2 Galligan JJ, Petersen DR (July 2012). "The human protein disulfide isomerase gene family". Human Genomics. 6 (1): 6. PMC 3500226
 . PMID 23245351. doi:10.1186/1479-7364-6-6.
. PMID 23245351. doi:10.1186/1479-7364-6-6. - 1 2 3 4 5 6 7 8 9 10 Perri, Emma R.; Thomas, Colleen J.; Parakh, Sonam; Spencer, Damian M.; Atkin, Julie D. (2016). "The Unfolded Protein Response and the Role of Protein Disulfide Isomerase in Neurodegeneration". Frontiers in Cell and Developmental Biology. 3. ISSN 2296-634X. PMC 4705227 . PMID 26779479. doi:10.3389/fcell.2015.00080.
- 1 2 3 Bechtel, Tyler J.; Weerapana, Eranthie (2017-03-01). "From structure to redox: The diverse functional roles of disulfides and implications in disease". PROTEOMICS. 17 (6): n/a–n/a. ISSN 1615-9861. PMC 5367942 . PMID 28044432. doi:10.1002/pmic.201600391.
- 1 2 3 Soares Moretti, Ana Iochabel; Martins Laurindo, Francisco Rafael (2017-03-01). "Protein disulfide isomerases: Redox connections in and out of the endoplasmic reticulum". Archives of Biochemistry and Biophysics. The Chemistry of Redox Signaling. 617: 106–119. doi:10.1016/j.abb.2016.11.007.
- ↑ Erdogan, Alican J.; Riemer, Jan (2017-01-01). "Mitochondrial disulfide relay and its substrates: mechanisms in health and disease". Cell and Tissue Research. 367 (1): 59–72. ISSN 0302-766X. doi:10.1007/s00441-016-2481-z.
- ↑ Manganas, Phanee; MacPherson, Lisa; Tokatlidis, Kostas (2017-01-01). "Oxidative protein biogenesis and redox regulation in the mitochondrial intermembrane space". Cell and Tissue Research. 367 (1): 43–57. ISSN 0302-766X. PMC 5203823 . PMID 27632163. doi:10.1007/s00441-016-2488-5.
- 1 2 3 Oka, Ojore B. V.; Yeoh, Hui Y.; Bulleid, Neil J. (2015-07-15). "Thiol-disulfide exchange between the PDI family of oxidoreductases negates the requirement for an oxidase or reductase for each enzyme". Biochemical Journal. 469 (2): 279–288. ISSN 0264-6021. PMID 25989104. doi:10.1042/bj20141423.
- ↑ Hatahet F, Ruddock LW (Oct 2007). "Substrate recognition by the protein disulfide isomerases". The FEBS Journal. 274 (20): 5223–34. PMID 17892489. doi:10.1111/j.1742-4658.2007.06058.x.
- ↑ Wittenberg G, Danon A (2008). "Disulfide bond formation in chloroplasts". Plant Science. 175 (4): 459–466. doi:10.1016/j.plantsci.2008.05.011.
- ↑ Ryser HJ, Flückiger R (Aug 2005). "Progress in targeting HIV-1 entry". Drug Discovery Today. 10 (16): 1085–94. PMID 16182193. doi:10.1016/S1359-6446(05)03550-6.
- 1 2 3 McBean, G J; López, M G; Wallner, F K (2017-06-01). "Redox-based therapeutics in neurodegenerative disease". British Journal of Pharmacology. 174 (12): 1750–1770. ISSN 1476-5381. PMC 5446580 . PMID 27477685. doi:10.1111/bph.13551.
- ↑ Lundström J, Holmgren A (Jun 1990). "Protein disulfide-isomerase is a substrate for thioredoxin reductase and has thioredoxin-like activity". The Journal of Biological Chemistry. 265 (16): 9114–20. PMID 2188973.
- ↑ Lyles MM, Gilbert HF (Jan 1991). "Catalysis of the oxidative folding of ribonuclease A by protein disulfide isomerase: dependence of the rate on the composition of the redox buffer". Biochemistry. 30 (3): 613–9. PMID 1988050. doi:10.1021/bi00217a004.
- ↑ Raturi A, Mutus B (Jul 2007). "Characterization of redox state and reductase activity of protein disulfide isomerase under different redox environments using a sensitive fluorescent assay". Free Radical Biology & Medicine. 43 (1): 62–70. PMID 17561094. doi:10.1016/j.freeradbiomed.2007.03.025.
- ↑ Hoffstrom BG, Kaplan A, Letso R, Schmid DC, Turmel RS, Lo GJ, Stockwell BR. "Inhibitors of protein disulfide isomerase suppress apoptosis induced by misfolded proteins" Nat". Chem. Biol. 2010 (12): 6. doi:10.1038/nchembio.467.
- ↑ Kaplan A, Gaschler MM, Dunn DE, Colligan R, Brown LM, Palmer AG, Lo DC, Stockwell BR (2015). "Small molecule induced oxidation of protein disulfide isomerase is neuroprotective". PNAS. 112: E2245–E2252. PMC 4418888 . PMID 25848045. doi:10.1073/pnas.1500439112.
- ↑ Galinski, CN; et al. (2016). "Revisiting the mechanistic basis of the French Paradox: Red wine inhibits the activity of protein disulfide isomerase in vitro". Thrombosis Research. 137: 169–173. PMID 26585763. doi:10.1016/j.thromres.2015.11.003.
- ↑ Ellgaard L, Ruddock LW (Jan 2005). "The human protein disulphide isomerase family: substrate interactions and functional properties". EMBO Reports. 6 (1): 28–32. PMC 1299221 . PMID 15643448. doi:10.1038/sj.embor.7400311.
- ↑ Appenzeller-Herzog C, Ellgaard L (Apr 2008). "The human PDI family: versatility packed into a single fold". Biochimica et Biophysica Acta. 1783 (4): 535–48. PMID 18093543. doi:10.1016/j.bbamcr.2007.11.010.
External links
- Protein Disulfide-Isomerase at the US National Library of Medicine Medical Subject Headings (MeSH)