Poikilotherm
| Thermoregulation in animals |
|---|
 |

A poikilotherm (/ˈpɔɪkələˌθɜːrm, pɔɪˈkɪləˌθɜːrm/) is an animal whose internal temperature varies considerably. It is the opposite of a homeotherm, an animal which maintains thermal homeostasis. While the term in principle can apply to all organisms, it is usually only applied to animals, and mostly to vertebrates. Usually the variation is a consequence of variation in the ambient environmental temperature. Many terrestrial ectotherms are poikilothermic.[1] However some ectotherms remain in temperature-constant environments to the point that they are actually able to maintain a constant internal temperature (i.e. are homeothermic). It is this distinction that often makes the term "poikilotherm" more useful than the vernacular "cold-blooded", which is sometimes used to refer to ectotherms more generally.
Poikilothermic animals include types of vertebrate animals, specifically fish, amphibians, and reptiles, as well as a large number of invertebrate animals. The naked mole-rat is the only mammal that is currently thought to be poikilothermic.[2][3]
Etymology
The term derives from Greek poikilos (ποικίλος), meaning "varied," ultimately from "thousand-making" and thermos (θερμός), meaning "heat".
Physiology
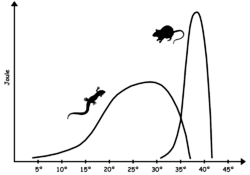
Poikilotherm animals must be able to function over a wider range of temperatures than homeotherms. The speed of most chemical reactions vary with temperature, and in order to function poikilotherms may have four to ten enzyme systems that operate at different temperatures for an important chemical reaction.[4] As a result, poikilotherms often have larger, more complex genomes than homeotherms in the same ecological niche. Frogs are a notable example of this effect, though their complex development is also an important factor in their large genome.[5]
Because their metabolism is variable and generally below that of homeothermic animals, sustained high-energy activities like powered flight in large animals or maintaining a large brain is generally beyond poikilotherm animals.[6] The metabolism of poikilotherms favors strategies such as sit-and-wait hunting over chasing prey for larger animals with high movement cost. As they do not use their metabolisms to heat or cool themselves, total energy requirement over time is low. For the same body weight, poikilotherms need only 5 to 10% of the energy of homeotherms.[7]
Adaptations in poikilotherms
- Some adaptations are behavioral. Lizards and snakes bask in the sun in the early morning and late evening, and seek shelter around noon.
- The eggs of the yellow-faced bumblebee are unable to regulate heat. A behavioral adaptation to combat this is incubation, where to maintain the internal temperatures of eggs, the queen and her workers will incubate the brood almost constantly, by warming their abdomens and touching them to the eggs. The bumblebee generates heat by shivering flight muscles even though they are not flying.
- Termite mounds are usually oriented in a north-south direction so that they absorb as much heat as possible around dawn and dusk and minimise heat absorption around noon.
- Tuna are able to warm their entire bodies through a heat exchange mechanism called the rete mirabile, which helps keep heat inside the body, and minimises the loss of heat through the gills. They also have their swimming muscles near the center of their bodies instead of near the surface, which minimises heat loss.
- Gigantothermy means using a low ratio of surface area to volume to minimise heat loss, such as in sea turtles.
- Camels, although they are homeotherms, thermoregulate using a method termed "temperature cycling" to conserve energy. In hot deserts, they allow their body temperature to rise during the day and fall during the night, adjusting their body temperature to cycle over approximately 6°C.[8]
Ecology
It is comparatively easy for a poikilotherm to accumulate enough energy to reproduce. Poikilotherms at the same trophic level often have much shorter generations than homeotherms: weeks rather than years. Such applies even to animals with similar ecological roles such as cats and snakes.
This difference in energy requirement also means that a given food source can support a greater density of poikilothermic animals than homeothermic animals.[9] This is reflected in the predator-prey ratio which is usually higher in poikilothermic fauna compared to homeothermic ones. However, when homeotherms and poikilotherms have similar niches, and compete, the homeotherm can often drive poikilothermic competitors to extinction, because homeotherms can gather food for a greater fraction of each day.
In medicine
In medicine, loss of normal thermoregulation in humans is referred to as "poikilothermia". This is usually seen with sedative and hypnotic drugs or in 'compartment syndrome'. For example, barbiturates, ethanol, and chloral hydrate may precipitate this effect. REM sleep is also considered a poikilothermic state in humans.[10]
Notes
- ↑ Milton Hildebrand; G. E. Goslow, Jr. Principal ill. Viola Hildebrand. (2001). Analysis of vertebrate structure. New York: Wiley. p. 429. ISBN 0-471-29505-1.
- ↑ Daly, T.J.M., Williams, L.A. and Buffenstein, R., (1997). Catecholaminergic innervation of interscapular brown adipose tissue in the naked mole-rat (Heterocephalus glaber). Journal of Anatomy, 190: 321-326. doi:10.1046/j.1469-7580.1997.19030321.x
- ↑ Sherwin, C.M. (2010). The Husbandry and Welfare of Non-traditional Laboratory Rodents. In "UFAW Handbook on the Care and Management of Laboratory Animals", R. Hubrecht and J. Kirkwood (Eds). Wiley-Blackwell. Chapter 25, pp. 359-369
- ↑ Cavalier-Smith, T. "Coevolution of vertebrate genome, cell, and nuclear sizes". Symposium on the Evolution of Terrestrial Vertebrates: 51–86.
- ↑ Ryan Gregory, T. (1 January 2002). "Genome size and developmental complexity". Genetica. 115 (1): 131–146. doi:10.1023/A:1016032400147.
- ↑ Willmer, P., Stone, G., & Johnston, I. A. (2000): Environmental physiology of animals. Blackwell Science, London. 644 pages, ISBN 0-632-03517-X.
- ↑ Campbell, N. A., Reece, J. B., et al. (2002). Biology. 6th edition. Benjamin / Cummings Publishing Company.
- ↑ Hill, Richard (2016). Animal Physiology. Sunderland, MA: Sinauer Associates. p. 270. ISBN 978-1605354712.
- ↑ Steen, J.B, Steen, H. & Stenseth, N.C. (1991): Population Dynamics of Poikilotherm and Homeotherm Vertebrates: Effects of Food Shortage. OICOS Vol. 60, No 2 (March, 1991), pp 269-272. summary
- ↑ Leon Rosenthal (2009). "3". In Teofilo Lee-Chiong. Sleep Medicine Essentials. Wiley-Blackwell. p. 12.
External links
 The dictionary definition of poikilotherm at Wiktionary
The dictionary definition of poikilotherm at Wiktionary