Onega stepanovi
| Onega stepanovi Temporal range: Ediacaran, around 555 Ma | |
|---|---|
 | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Proarticulata |
| Genus: | Onega |
| Species: | O. stepanovi |
| Binomial name | |
| Onega stepanovi Fedonkin, 1976[1] | |
Onega stepanovi is a fossil organism from Ediacaran deposites of the Arkhangelsk Region, Russia. It was described by Mikhail A. Fedonkin in 1976[1]
Etymology
The generic name Onega comes from the Onega Peninsula of the White Sea, where the first fossils were found. The specific name is given in honour of V. A. Stepanov, discoverer of the first Ediacaran fossil locality in Arkhangelsk Region, on the Letniy Bereg (Summer Coast) of the Onega Peninsula in 1972.[2]
Morphology
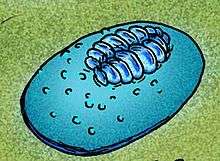
The small fossils, which range up to 7 millimetres (0.28 in) long, have oval outlines and low bodies with an articulated central zone built of isomers encircled by an undivided zone. The surface of the undivided region of Onega is covered with small tubercles.[3]
Onega was originally described by Mikhail Fedonkin as a problematic organism, being grouped together with Vendia, Praecambridium and Vendomia as possible stem-group arthropods due to a vague similarity with primitive Cambrian trilobites and arthropods.[1]
In 1985 Mikhail Fedonkin erected Phylum Proarticulata,[4] in which he placed: Onega, Dickinsonia, Palaeoplatoda, Vendia, Vendomia, Praecambridium and Pseudovendia sp., although he did not exclude the possibility that Onega may still be related to various lower Cambrian arthropods, such as Skania.[4][5]
Andrey Yu. Ivantsov has proposed that Onega be placed in phylum Proarticulata, as the segments in recently discovered, exceptionally well-preserved fossils display the glide, or "staggered", symmetry characteristic of the majority of proarticulatans.[3]
Fossil record
Imprints of Onega stepanovi have been found in the Verkhovka and Zimnie Gory Formations[6][7] of the Ediacaran (Vendian) rocks of the Arkhangelsk Region, Russia.[3]
All the fossil specimens are negative imprints on the bases of fine-grained sandstone beds with the "elephant skin" and tubercle texture diagnostic of microbial mats.[8] The same bedding planes contain various other Ediacaran species: Cyclomedusa, Ediacaria, palaeopascichnids, Eoporpita, Yorgia, Andiva, Archaeaspinus, Vendia, Dickinsonia, Anfesta, Albumares, Tribrachidium, Kimberella, Parvancorina, Charniodiscus and others.
See also
References
- 1 2 3 B. M. Keller & M. A. Fedonkin (1976). Новые находки окаменелостей в валдайской серии докембрия по р. Сюзьме [New Records of Fossils in the Valdaian Group of the Precambrian on the Syuz'ma River] (PDF). Izv. Akad. Nauk SSSR, Ser. Geol. (in Russian). 3: 38–44.
- ↑ Keller, B. M.; Menner, V. V.; Stepanov, V. A. & Chumakov, N. M. (1974). "Новые находки Metazoa в вендомии Руссой платформы" [New Finds of Metazoa in the Vendomii of the Russian Platform] (PDF). Izv. Akad. Nauk SSSR, Ser. Geol. (in Russian). 12: 130–134.
- 1 2 3 Ivantsov, A. Yu. (April 2007). "Small Vendian transversely Articulated fossils". Paleontological Journal. 41 (2): 113. doi:10.1134/S0031030107020013.
- 1 2 M. A. Fedonkin (1985). "Systematic Description of Vendian Metazoa". Vendian System: Historical–Geological and Paleontological Foundation, Vol. 1: Paleontology. Moscow: Nauka, pp. 70–106.
- ↑ Stefan Hengtson; Mikhail A. Fedonkin; Jere H. Lipps. (1992). "The Major Biotas of Proterozoic to Early Cambrian Multicellular Organisms". In: Edited by J. William Schopf; Cornelis Klein. Proterozoic Biosphere. Cambridge University Press, pp. 433–435.
- ↑ D. V. Grazhdankin (2003). "The Structure and Sedimentation Conditions in the Vendian assemblage in the Southeastern White Sea" (PDF). Stratigraphy and Geological Correlation. 11 (4): 313–331.
- ↑ D. V. Grazhdankin (2004). "Patterns of distribution in the Ediacaran biotas: facies versus biogeography and evolution" (PDF). Paleobiology. 30 (2): 203–221. doi:10.1666/0094-8373(2004)030<0203:PODITE>2.0.CO;2.
- ↑ Gehling, J. G. (1999). "Microbial mats in terminal Proterozoic siliciclastics: Ediacaran death masks". PALAIOS. 14 (1): 40–57. doi:10.2307/3515360.