Myelin
| Myelin sheath |
|---|
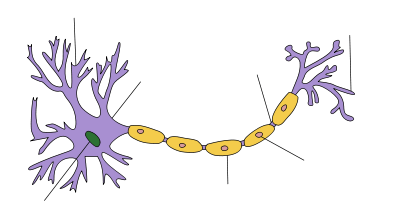
Myelin is a fatty white substance that surrounds the axon of some nerve cells, forming an electrically insulating layer. It is essential for the proper functioning of the nervous system. It is an outgrowth of a type of glial cell.
The production of the myelin sheath is called myelination or myelinogenesis. In humans, myelination begins early in the 3rd trimester,[1] although little myelin exists in the brain at the time of birth. During infancy, myelination occurs quickly, leading to a child's fast development, including crawling and walking in the first year. Myelination continues through the adolescent stage of life.
Schwann cells supply the myelin for the peripheral nervous system, whereas oligodendrocytes, specifically of the interfascicular type, myelinate the axons of the central nervous system. Myelin is considered a defining characteristic of the (gnathostome) vertebrates, but myelin-like sheaths have also been seen in some invertebrates, although they are quite different from vertebrate myelin at the molecular level. Myelin was discovered in 1854 by Rudolf Virchow.[2]
Composition

Myelin comprises different cell types and varies in chemical composition and configuration but performs the same insulating function. Myelinated axons are white; hence, the "white matter" of the brain. Myelin insulates axons from electrically charged atoms and molecules. These charged particles (ions) are found in the fluid surrounding the entire nervous system. Under a microscope, myelin looks like strings of sausages.
Cholesterol is an essential component of myelin,[3] which comprises about 40% water; the dry mass comprises between 70% and 85% lipids and between 15% and 30% proteins. Myelin basic protein (MBP) constitutes ~30% of myelin protein,[4] myelin oligodendrocyte glycoprotein, and proteolipid protein (PLP, which makes up ~50% of myelin protein[5]). The primary lipid of myelin is a glycolipid called galactocerebroside. The intertwining hydrocarbon chains of sphingomyelin strengthen the myelin sheath. In brain, the myelin sheath covers the fibers of the corpus callosum, which constitute the inner part of the cerebral hemisphere.
Function

.svg.png)
1. Axon
2. Nucleus of Schwann Cell
3. Schwann Cell
4. Myelin Sheath
5. Neurilemma
The main purpose of a myelin sheath is to increase the speed at which impulses propagate along the myelinated fiber. Along unmyelinated fibers, impulses continuously move as waves, but, in myelinated fibers, they "hop" or propagate by saltatory conduction. Myelin decreases capacitance and increases electrical resistance across the cell membrane (the axolemma). Thus, myelination prevents the electric current from leaving the axon. It has been suggested that myelin permits larger body size by maintaining agile communication between distant body parts.[6]
Myelinated fibers lack voltage-gated ion channels (approximately 25 μm−2) along the myelinated internodes, exposing them only at the nodes of Ranvier. Here, they are found far more abundantly (between 2,000 and 12,000 μm−2).[7] Myelinated fibers succeed in reducing sodium leakage into the extracellular fluid (ECF), maintaining a strong separation of charge between the intracellular fluid (ICF) and the ECF. This increases sodium’s ability to travel along the axon more freely. However, the sodium diffuses along the axolemma rapidly, but is decremental by nature. The sodium cannot trigger the opening of the voltage-gated sodium channels as it becomes weaker. The nodes of Ranvier, being exposed to the ECF every 1 mm or so, contain large amounts of voltage-gated sodium channels, and allow enough sodium into the axon to regenerate the action potential.[8] Each time the action potential reaches a node of Ranvier, it is restored to its original action potential (+35 mV).[7]
When a peripheral fiber is severed, the myelin sheath provides a track along which regrowth can occur. However, the myelin layer does not ensure a perfect regeneration of the nerve fiber. Some regenerated nerve fibers do not find the correct muscle fibers, and some damaged motor neurons of the peripheral nervous system die without regrowth. Damage to the myelin sheath and nerve fiber is often associated with increased functional insufficiency.
Unmyelinated fibers and myelinated axons of the mammalian central nervous system do not regenerate.
Some studies have revealed that optic nerve fibers can be regenerated in postnatal rats. This regeneration depends upon two conditions: axonal die-back has to be prevented with appropriate neurotrophic factors, and neurite growth inhibitory components have to be inactivated. These studies may lead to further understanding of nerve fiber regeneration in the central nervous system.
Disorders
Demyelination
Demyelination is the loss of the myelin sheath insulating the nerves, and is the hallmark of some neurodegenerative autoimmune diseases, including multiple sclerosis, acute disseminated encephalomyelitis, neuromyelitis optica, transverse myelitis, chronic inflammatory demyelinating polyneuropathy, Guillain–Barré syndrome, central pontine myelinosis, inherited demyelinating diseases such as leukodystrophy, and Charcot-Marie-Tooth disease. Sufferers of pernicious anaemia can also suffer nerve damage if the condition is not diagnosed quickly. Subacute combined degeneration of spinal cord secondary to pernicious anaemia can lead to slight peripheral nerve damage to severe damage to the central nervous system, affecting speech, balance, and cognitive awareness. When myelin degrades, conduction of signals along the nerve can be impaired or lost, and the nerve eventually withers. A more serious case of myelin deterioration is called Canavan disease.
The immune system may play a role in demyelination associated with such diseases, including inflammation causing demyelination by overproduction of cytokines via upregulation of tumor necrosis factor[9] or interferon.
Symptoms
Demyelination results in diverse symptoms determined by the functions of the affected neurons. It disrupts signals between the brain and other parts of the body; symptoms differ from patient to patient, and have different presentations upon clinical observation and in laboratory studies.
Typical symptoms include:
- blurriness in the central visual field that affects only one eye, may be accompanied by pain upon eye movement
- double vision
- loss of vision/hearing
- odd sensation in legs, arms, chest, or face, such as tingling or numbness (neuropathy)
- weakness of arms or legs
- cognitive disruption, including speech impairment and memory loss
- heat sensitivity (symptoms worsen or reappear upon exposure to heat, such as a hot shower)
- loss of dexterity
- difficulty coordinating movement or balance disorder
- difficulty controlling bowel movements or urination
- fatigue
- tinnitus[10]
Myelin repair
Research to repair damaged myelin sheaths is ongoing. Techniques include surgically implanting oligodendrocyte precursor cells in the central nervous system and inducing myelin repair with certain antibodies. While results in mice have been encouraging (via stem cell transplantation), whether this technique can be effective in replacing myelin loss in humans is still unknown.[11] Cholinergic treatments, such as acetylcholinesterase inhibitors (AChEIs), may have beneficial effects on myelination, myelin repair, and myelin integrity. Increasing cholinergic stimulation also may act through subtle trophic effects on brain developmental processes and particularly on oligodendrocytes and the lifelong myelination process they support. By increasing oligodendrocyte cholinergic stimulation, AChEIs, and other cholinergic treatments, such as nicotine, possibly could promote myelination during development and myelin repair in older age.[12] Glycogen synthase kinase 3β inhibitors such as lithium chloride have been found to promote myelination in mice with damaged facial nerves.[13] Cholesterol is a necessary nutrient for the myelin sheath.
Dysmyelination
Dysmyelination is characterized by a defective structure and function of myelin sheaths; unlike demyelination, it does not produce lesions. Such defective sheaths often arise from genetic mutations affecting the biosynthesis and formation of myelin. The shiverer mouse represents one animal model of dysmyelination. Human diseases where dysmyelination has been implicated include leukodystrophies (Pelizaeus–Merzbacher disease, Canavan disease, phenylketonuria) and schizophrenia.[14][15][16]
Invertebrate myelin
Functionally equivalent myelin-like sheaths are found in several invertebrate taxa including Oligochaete, Penaeid, Palaemonid, and Calanoids. These myelin-like sheaths share several structural features with the sheaths found in vertebrates including multiplicity of membranes, condensation of membrane, and nodes.[6] However, the nodes in vertebrates are annular; i.e., they encircle the axon. In contrast, nodes found in the sheaths of invertebrates are either annular or fenestrated; i.e., they are restricted to "spots." It is notable that the fastest recorded conduction speed (across both vertebrates and invertebrates) is found in the ensheathed axons of the Kuruma shrimp, an invertebrate.[6]
See also
- The Myelin Project, project to regenerate myelin
- Myelin Repair Foundation, a nonprofit medical research foundation for multiple sclerosis drug discovery.
- Myelin protein zero
References
- ↑ "Pediatric Neurologic Examination Videos & Descriptions: Developmental Anatomy". library.med.utah.edu. Retrieved 2016-08-20.
- ↑ Virchow R (1854). "Über das ausgebreitete Vorkommen einer dem Nervenmark analogen Substanz in den tierischen Geweben". Archiv für pathologische Anatomie und Physiologie und für klinische Medizin (in German). 6 (4): 562–72. doi:10.1007/BF02116709.
- ↑ Gesine Saher; Britta Brügger; Corinna Lappe-Siefke; Wiebke Möbius; Ryu-ichi Tozawa; Michael C Wehr; Felix Wieland; Shun Ishibashi; Klaus-Armin Nave (March 2005). "High cholesterol level is essential for myelin membrane growth". Nature Neuroscience. 8: 468–475. doi:10.1038/nn1426.
- ↑ Steinman, Lawrence (May 3, 1996). "Multiple Sclerosis: A Coordinated Immunological Attack against Myelin in the Central Nervous System". Cell. 85: 299–302. PMID 8616884. doi:10.1016/S0092-8674(00)81107-1. Retrieved 2016-02-03.
- ↑ Mallucci, Giulia; Peruzzotti-Jametti, Luca; Bernstock, Joshua D.; Pluchino, Stefano (2015-04-01). "The role of immune cells, glia and neurons in white and gray matter pathology in multiple sclerosis". Progress in Neurobiology. 127–128: 1–22. PMC 4578232
 . PMID 25802011. doi:10.1016/j.pneurobio.2015.02.003.
. PMID 25802011. doi:10.1016/j.pneurobio.2015.02.003. - 1 2 3 Hartline Daniel K (2008). "What is myelin?.". Neuron Glia Biology. 4: 153–163. doi:10.1017/S1740925X09990263.
- 1 2 Saladin, Kenneth S.. Anatomy & physiology: the unity of form and function. 6th ed. New York, NY: McGraw-Hill, 2012. Print.
- ↑ Siegel GJ, Agranoff BW, Albers RW, et al., editors. Basic Neurochemistry: Molecular, Cellular and Medical Aspects. 6th edition. Philadelphia: Lippincott-Raven; 1999. Available from: http://www.ncbi.nlm.nih.gov/books/NBK20385/
- ↑ Ledeen RW, Chakraborty G (March 1998). "Cytokines, signal transduction, and inflammatory demyelination: review and hypothesis". Neurochem. Res. 23 (3): 277–89. PMID 9482240. doi:10.1023/A:1022493013904.
- ↑ Mayo Clinic 2007 and University of Leicester Clinical Studies, 2014
- ↑ Windrem MS, Nunes MC, Rashbaum WK, Schwartz TH, Goodman RA, McKhann G, Roy NS, Goldman SA (January 2004). "Fetal and adult human oligodendrocyte progenitor cell isolates myelinate the congenitally dysmyelinated brain". Nat. Med. 10 (1): 93–7. PMID 14702638. doi:10.1038/nm974. Lay summary – www.futurepundit.com.
- ↑ Bartzokis G (August 2007). "Acetylcholinesterase inhibitors may improve myelin integrity". Biol. Psychiatry. 62 (4): 294–301. PMID 17070782. doi:10.1016/j.biopsych.2006.08.020.
- ↑ Makoukji J, Belle M, Meffre D, Stassart R, Grenier J, Shackleford G, Fledrich R, Fonte C, Branchu J, Goulard M, de Waele C, Charbonnier F, Sereda MW, Baulieu EE, Schumacher M, Bernard S, Massaad C (March 2012). "Lithium enhances remyelination of peripheral nerves". Proc. Natl. Acad. Sci. U.S.A. 109 (10): 3973–8. PMC 3309729 . PMID 22355115. doi:10.1073/pnas.1121367109.
- ↑ Krämer-Albers EM, Gehrig-Burger K, Thiele C, Trotter J, Nave KA (November 2006). "Perturbed interactions of mutant proteolipid protein/DM20 with cholesterol and lipid rafts in oligodendroglia: implications for dysmyelination in spastic paraplegia". J. Neurosci. 26 (45): 11743–52. PMID 17093095. doi:10.1523/JNEUROSCI.3581-06.2006.
- ↑ Matalon R, Michals-Matalon K, Surendran S, Tyring SK (2006). "Canavan disease: studies on the knockout mouse". Adv. Exp. Med. Biol. Advances in Experimental Medicine and Biology. 576: 77–93; discussion 361–3. ISBN 978-0-387-30171-6. PMID 16802706. doi:10.1007/0-387-30172-0_6.
- ↑ Tkachev D, Mimmack ML, Huffaker SJ, Ryan M, Bahn S (August 2007). "Further evidence for altered myelin biosynthesis and glutamatergic dysfunction in schizophrenia". Int. J. Neuropsychopharmacol. 10 (4): 557–63. PMID 17291371. doi:10.1017/S1461145706007334.
External links
- The Myelin Project
- Athabasca University Biological Psychology Website
- The MS Information Sourcebook, Myelin
- The Myelin Repair Foundation
- H & E Histology
- Luxol Fast Blue: Modified Kluver's Method to stain for Myelin Sheath
- Radiology and Pathology of Myelin the MedPix Medical Image Database
- Archives of Neurology
- American Society for Neurochemistry