MMP1





Matrix metalloproteinase-1 (MMP-1) also known as interstitial collagenase and fibroblast collagenase is an enzyme that in humans is encoded by the MMP1 gene.[5][6][7] The gene is part of a cluster of MMP genes which localize to chromosome 11q22.3.[5] MMP-1 was the first vertebrate collagenase both purified to homogeneity as a protein, and cloned as a cDNA.[8][9]
Structural Features
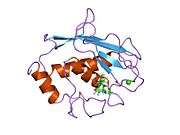
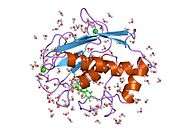
MMP-1 has an archetypal structure consisting of a pre-domain, a pro-domain, a catalytic domain, a linker region and a hemopexin-like domain.[10] The primary structure of MMP-1 was first published by Goldberg, G I, et al.[9] Two main nomenclatures for the primary structure are currently in use, the original one from which the first amino-acid starts with the signalling peptide and a second one where the first amino-acid starts counting from the prodomain (proenzyme nomenclature).
Catalytic Domain
The Catalytic Domains of MMPs share very similar characteristics, having a general shape of oblate ellipsoid with a diameter of ~40 Å.[11] Despite the similarity of the Catalytic domains of MMPs, this entry will focus only on the structural features of MMP-1 Catalytic Domain.
Overall Structural Characteristics
The Catalytic Domain of MMP-1 is composed of five highly twisted β-strands (sI-sV), three α-helix (hA-hC) and a total of eight loops, enclosing a total of five metal ions, three Ca2+ and two Zn2+, one of which with catalytic role.[12]
The Catalytic Domain (CAT) of MMP-1 starts with the F100 (non-truncated CAT) as the first amino-acid of the N-terminal loop of the CAT domain. The first published x-ray structure of the CAT domain was representative of the truncated form of this domain, where the first 7 amino-acids are not present.[12]
After the initial loop, the sequences follows to the first and longest β-sheet (sI). A second loop precedes large "amphipathic α-helix" (hA) that longitudinally spans protein site. The β-strands sII and sIII follows separated by the respective loops, loop 4 being commonly designated as "short loop" bridging sII to sIII. Following the sIII strand the sequence meets the 'S-shaped double loop' that is of primary importance for the peptide structure and catalytic activity (see further) as it extends to the cleft side "bulge", continuing to the only antiparallel β-strand sIV, which is prime importance for binding peptidic substrates or inhibitors by forming main chain hydrogen bond. Following sIV, loop Gln186-Gly192 and β-strand sV are responsible for contributing with many ligands to the several metal ions present in the protein (read further). A large open loop follows sV which has proven importance in substrate specificity within the MMPs family.[13] A specific region (183)RWTNNFREY(191) as been identified as a critical segment of matrix metalloproteinase 1 for the expression of collagenolytic activity.[14] On C-terminal part of the CAT Domain the hB α-helix, known as the "active-site helix" encompasses part of the "zinc-binding consensus sequence" HEXXHXXGXXH that is characteristic of the Metzincin superfamily.[15][16] The α-helix hB finishes abruptly at Gly225 where the last loop of the domain starts. This last loop contains the "specificity loop" which is the shortest in the MMPs family. The Catalytic Domain ends at Gly261 with α-helix hC.
Function
MMPs are involved in the breakdown of extracellular matrix in normal physiological processes, such as embryonic development, reproduction, and tissue remodeling, as well as in disease processes, such as arthritis and metastasis. Specifically, MMP-1 breaks down the interstitial collagens, types I, II, and III.
 Induction of matrix metalloproteinase 1 in rat corneas by ciprofloxacin, ofloxacin and levofloxacin (b,c,d) compared to artificial tears (a). Reviglio et al., 2003. |
Regulation
Mechanical force may increase the expression of MMP1 in human periodontal ligament cells.[17]
Interactions
MMP1 has been shown to interact with CD49b.[18][19]
References
- 1 2 3 GRCh38: Ensembl release 89: ENSG00000196611 - Ensembl, May 2017
- 1 2 3 GRCm38: Ensembl release 89: ENSMUSG00000043089 - Ensembl, May 2017
- ↑ "Human PubMed Reference:".
- ↑ "Mouse PubMed Reference:".
- 1 2 EntrezGene 4312
- ↑ Brinckerhoff CE, Ruby PL, Austin SD, Fini ME, White HD (February 1987). "Molecular cloning of human synovial cell collagenase and selection of a single gene from genomic DNA". J. Clin. Invest. 79 (2): 542–6. PMC 424122
 . PMID 3027129. doi:10.1172/JCI112845.
. PMID 3027129. doi:10.1172/JCI112845. - ↑ Pendás AM, Santamaría I, Alvarez MV, Pritchard M, López-Otín C (October 1996). "Fine physical mapping of the human matrix metalloproteinase genes clustered on chromosome 11q22.3". Genomics. 37 (2): 266–8. PMID 8921407. doi:10.1006/geno.1996.0557.
- ↑ Gross J, Lapiere CM (June 1962). "Collagenolytic activity in amphibian tissues: a tissue culture assay". Proc. Natl. Acad. Sci. U.S.A. 48 (6): 1014–22. PMC 220898 . PMID 13902219. doi:10.1073/pnas.48.6.1014.
- 1 2 Goldberg GI, Wilhelm SM, Kronberger A, Bauer EA, Grant GA, Eisen AZ (May 1986). "Human fibroblast collagenase. Complete primary structure and homology to an oncogene transformation-induced rat protein". J. Biol. Chem. 261 (14): 6600–5. PMID 3009463.
- ↑ Li J, Brick P, O'Hare MC, Skarzynski T, Lloyd LF, Curry VA, Clark IM, Bigg HF, Hazleman BL, Cawston TE (June 1995). "Structure of full-length porcine synovial collagenase reveals a C-terminal domain containing a calcium-linked, four-bladed beta-propeller". Structure. 3 (6): 541–9. PMID 8590015. doi:10.1016/S0969-2126(01)00188-5.
- ↑ Tallant C, Marrero A, Gomis-Rüth FX (January 2010). "Matrix metalloproteinases: fold and function of their catalytic domains". Biochim. Biophys. Acta. 1803 (1): 20–8. PMID 19374923. doi:10.1016/j.bbamcr.2009.04.003.
- 1 2 Spurlino JC, Smallwood AM, Carlton DD, Banks TM, Vavra KJ, Johnson JS, Cook ER, Falvo J, Wahl RC, Pulvino TA (June 1994). "1.56 A structure of mature truncated human fibroblast collagenase". Proteins. 19 (2): 98–109. PMID 8090713. doi:10.1002/prot.340190203.
- ↑ Maskos K, Bode W (November 2003). "Structural basis of matrix metalloproteinases and tissue inhibitors of metalloproteinases". Mol. Biotechnol. 25 (3): 241–66. PMID 14668538. doi:10.1385/MB:25:3:241.
- ↑ Chung L, Shimokawa K, Dinakarpandian D, Grams F, Fields GB, Nagase H (September 2000). "Identification of the (183)RWTNNFREY(191) region as a critical segment of matrix metalloproteinase 1 for the expression of collagenolytic activity". J. Biol. Chem. 275 (38): 29610–7. PMID 10871619. doi:10.1074/jbc.M004039200.
- ↑ Bode W, Gomis-Rüth FX, Stöckler W (September 1993). "Astacins, serralysins, snake venom and matrix metalloproteinases exhibit identical zinc-binding environments (HEXXHXXGXXH and Met-turn) and topologies and should be grouped into a common family, the 'metzincins'". FEBS Lett. 331 (1-2): 134–40. PMID 8405391. doi:10.1016/0014-5793(93)80312-I.
- ↑ Stöcker W, Grams F, Baumann U, Reinemer P, Gomis-Rüth FX, McKay DB, Bode W (May 1995). "The metzincins--topological and sequential relations between the astacins, adamalysins, serralysins, and matrixins (collagenases) define a superfamily of zinc-peptidases". Protein Sci. 4 (5): 823–40. PMC 2143131 . PMID 7663339. doi:10.1002/pro.5560040502.
- ↑ Huang SF, Li YH, Ren YJ, Cao ZG, Long X (August 2008). "The effect of a single nucleotide polymorphism in the matrix metalloproteinase-1 (MMP-1) promoter on force-induced MMP-1 expression in human periodontal ligament cells". Eur. J. Oral Sci. 116 (4): 319–23. PMID 18705799. doi:10.1111/j.1600-0722.2008.00552.x.
- ↑ Stricker TP, Dumin JA, Dickeson SK, Chung L, Nagase H, Parks WC, Santoro SA (August 2001). "Structural analysis of the alpha(2) integrin I domain/procollagenase-1 (matrix metalloproteinase-1) interaction". J. Biol. Chem. 276 (31): 29375–81. PMID 11359774. doi:10.1074/jbc.M102217200.
- ↑ Dumin JA, Dickeson SK, Stricker TP, Bhattacharyya-Pakrasi M, Roby JD, Santoro SA, Parks WC (August 2001). "Pro-collagenase-1 (matrix metalloproteinase-1) binds the alpha(2)beta(1) integrin upon release from keratinocytes migrating on type I collagen". J. Biol. Chem. 276 (31): 29368–74. PMID 11359786. doi:10.1074/jbc.M104179200.
Further reading
- Krane SM (1995). "Is collagenase (matrix metalloproteinase-1) necessary for bone and other connective tissue remodeling?". Clin. Orthop. Relat. Res. (313): 47–53. PMID 7641497.
- Massova I, Kotra LP, Fridman R, Mobashery S (1998). "Matrix metalloproteinases: structures, evolution, and diversification". FASEB J. 12 (12): 1075–95. PMID 9737711. doi:10.1142/S0217984998001256.
- Nagase H, Woessner JF (1999). "Matrix metalloproteinases". J. Biol. Chem. 274 (31): 21491–4. PMID 10419448. doi:10.1074/jbc.274.31.21491.
- Okada Y, Hashimoto G (2001). "[Degradation of extracellular matrix by matrix metalloproteinases and joint destruction]". Seikagaku. 73 (11): 1309–21. PMID 11831026.
- Seiki M (2003). "Membrane-type 1 matrix metalloproteinase: a key enzyme for tumor invasion". Cancer Lett. 194 (1): 1–11. PMID 12706853. doi:10.1016/S0304-3835(02)00699-7.
- Golubkov VS, Strongin AY (2007). "Proteolysis-driven oncogenesis". Cell Cycle. 6 (2): 147–50. PMID 17245132. doi:10.4161/cc.6.2.3706.
External links
PDB gallery | ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||