Isocitrate lyase
| Isocitrate Lyase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Homotetrameric structure of Isocitrate lyase from E. coli. Based on PDB 1IGW.[1] | |||||||||
| Identifiers | |||||||||
| EC number | 4.1.3.1 | ||||||||
| CAS number | 9045-78-7 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / EGO | ||||||||
| |||||||||
| Isocitrate lyase family | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| Symbol | ICL | ||||||||
| Pfam | PF00463 | ||||||||
| InterPro | IPR000918 | ||||||||
| PROSITE | PDOC00145 | ||||||||
| SCOP | 1f8m | ||||||||
| SUPERFAMILY | 1f8m | ||||||||
| |||||||||
Isocitrate lyase (EC 4.1.3.1), or ICL, is an enzyme in the glyoxylate cycle that catalyzes the cleavage of isocitrate to succinate and glyoxylate.[2][3] Together with malate synthase, it bypasses the two decarboxylation steps of the tricarboxylic acid cycle (TCA cycle) and is used by bacteria, fungi, and plants.[4]
The systematic name of this enzyme class is isocitrate glyoxylate-lyase (succinate-forming). Other names in common use include isocitrase, isocitritase, isocitratase, threo-Ds-isocitrate glyoxylate-lyase, and isocitrate glyoxylate-lyase. This enzyme participates in glyoxylate and dicarboxylate metabolism.
Mechanism
This enzyme belongs to the family of lyases, specifically the oxo-acid-lyases, which cleave carbon-carbon bonds. Other enzymes also belong to this family including carboxyvinyl-carboxyphosphonate phosphorylmutase (EC 2.7.8.23) which catalyses the conversion of 1-carboxyvinyl carboxyphosphonate to 3-(hydrohydroxyphosphoryl) pyruvate carbon dioxide, and phosphoenolpyruvate mutase (EC 5.4.2.9), which is involved in the biosynthesis of phosphinothricin tripeptide antibiotics.
During catalysis, isocitrate is deprotonated, and an aldol cleavage results in the release of succinate and glyoxylate. This reaction mechanism functions much like that of aldolase in glycolysis, where a carbon-carbon bond is cleaved and an aldehyde is released.[5]
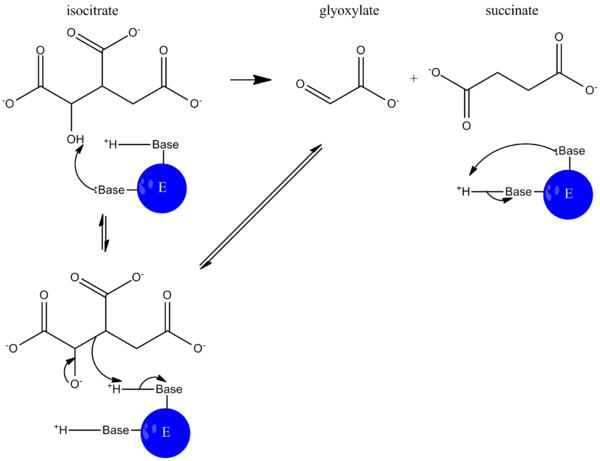
In the glyoxylate cycle, malate synthase then catalyzes the condensation of glyoxylate and acetyl-CoA to form malate so the cycle can continue.
ICL competes with isocitrate dehydrogenase, an enzyme found in the TCA cycle, for isocitrate processing. Flux through these enzymes is controlled by phosphorylation of isocitrate dehydrogenase, which has a much higher affinity for isocitrate as compared to ICL.[6] Deactivation of isocitrate dehydrogenase by phosphorylation thus leads to increased isocitrate channeling through ICL, as seen when bacteria are grown on acetate, a two-carbon compound.[6]
Enzyme structure
As of late 2007, 5 structures have been solved for this class of enzymes, with PDB accession codes 1DQU, 1F61, 1F8I, 1F8M, and 1IGW.
ICL is composed of four identical chains and requires a Mg2+ or Mn2+ and a thiol for activity.[4] In Escherichia coli, Lys-193, Lys-194, Cys-195, His-197, and His-356 are thought to be catalytic residues, while His-184 is thought to be involved in the assembly of the tetrameric enzyme.[7]
Between prokaryotes and eukaryotes, a difference in ICL structure is the addition of approximately 100 amino acids near the center of the eukaryotic enzyme. In eukaryotes, the additional amino acids are thought to function in the localization of ICL to single-membrane-bound organelles called glyoxysomes.[4][8] These additional amino acids account for the difference in molecular mass: the prokaryotic ICL is 48kDa, while the eukaryotic ICL is 67 kDa.[4] Only one cysteine residue is conserved between the sequences of the fungal, plant and bacterial enzymes; it is located in the middle of a conserved hexapeptide.
Biological function
The ICL enzyme has been found to be functional in various archaea, bacteria, protists, plants, fungi, and nematodes.[9] Although the gene has been found in genomes of nematodes and cnidaria, it has not been found in the genomes of placental mammals.[9]
By diverting isocitrate from the TCA cycle, the actions of ICL and malate synthase in the glyoxylate cycle result in the net assimilation of carbon from 2-carbon compounds.[10] Thus, while the TCA cycle yields no net carbon assimilation, the glyoxylate cycle generates intermediates that can be used to synthesize glucose (via gluconeogenesis), plus other biosynthetic products. As a result, organisms that use ICL and malate synthase are able to synthesize glucose and its metabolic intermediates from acetyl-CoA derived from acetate or from the degradation of ethanol, fatty acids, or poly-β-hydroxybutyrate.[4]
This function is especially important for higher plants when using seed oils. In germinating seeds, the breakdown of oils generates acetyl-CoA. This serves as a substrate for the glyoxylate cycle, which generates intermediates which serve as a primary nutrient source prior to the beginning of production of sugars by photosynthesis.[8]
Disease relevance
ICL has found to be important in human, animal, and plant pathogenesis.[4] For several agricultural crops including cereals, cucumbers, and melons, increased expression of the gene encoding ICL is important for fungal virulence.[4] For instance, increased gene expression of icl1 has been seen in the fungus Leptosphaeria maculans upon infection of canola. Inactivation of the icl1 gene leads to reduced pathogenicity of the fungus, which is thought to be a result of the inability of the fungus to use carbon sources provided by the plant.[11]
Additionally, upregulation of the glyoxylate cycle has been seen for pathogens that attack humans. This is the case for fungi such as Candida albicans, which inhabits the skin, mouth, GI tract, gut and vagina of mammals and can lead to systemic infections of immunocompromised patients; as well as for the bacterium Mycobacterium tuberculosis, the major causative agent of tuberculosis.[12][13] In this latter case, ICL has been found to be essential for survival in the host.[14]
Because of its use by pathogenic fungi and bacteria, specific inhibitors are being sought for ICL and malate synthase.[4] Although some inhibitors have already been identified, including itaconate, itaconic anhydride, bromopyruvate, nitropropionate, oxalate, and malate, these are non-specific and would also inhibit other enzymes essential for host function.[4] More research is needed to identify inhibitors that selectively target enzymes in the glyoxylate cycle.
See also
References
- ↑ Britton, KL; Abeysinghe IS; Baker PJ; Barynin V; Diehl P; Langridge SJ; McFadden BA; Sedelnikova SE; Stillman TJ; Weeradechapon K; Rice DW (Sep 2001). "The structure and domain organization of Escherichia coli isocitrate lyase". Acta Crystallogr D. 57 (9): 1209–1218. PMID 11526312. doi:10.1107/S0907444901008642.
- ↑ Beeching JR (1989). "High sequence conservation between isocitrate lyase from Escherichia coli and Ricinus communis". Protein Seq. Data Anal. 2 (6): 463–466. PMID 2696959.
- ↑ Tanaka A, Atomi H, Ueda M, Hikida M, Hishida T, Teranishi Y (1990). "Peroxisomal isocitrate lyase of the n-alkane-assimilating yeast Candida tropicalis: gene analysis and characterization". J. Biochem. 107 (2): 262–266. PMID 2361956.
- 1 2 3 4 5 6 7 8 9 Dunn, MF; Ramirez-Trujill JA; Hernandez-Lucas I (Oct 2009). "Major roles of isocitrate lyase and malate synthase in bacterial and fungal pathogenesis". Microbiology. 155 (10): 3166–3175. PMID 19684068. doi:10.1099/mic.0.030858-0.
- ↑ Garrett R, Grisham CN (2008). Biochemistry. Brooks Cole. p. 588. ISBN 978-0-495-10935-8.
- 1 2 Cozzone, AJ (1998). "Regulation of acetate metabolism by protein phosphorylation in enteric bacteria". Annu. Rev. Microbiol. 52: 127–164. PMID 9891796. doi:10.1146/annurev.micro.52.1.127.
- ↑ Rehman, A; McFadden BA (Jul 1997). "Lysine 194 is functional in isocitrate lyase from Escherichia coli". Curr. Microbiol. 35 (1): 14–17. PMID 9175553. doi:10.1007/s002849900203.
- 1 2 Eastmond, PJ; Graham IA (Feb 2001). "Re-examining the role of the glyoxylate cycle in oilseeds". Trends Plant Sci. 6 (2): 72–78. PMID 11173291. doi:10.1016/S1360-1385(00)01835-5.
- 1 2 Kondrashov, FA; Koonin EV; Morgunov IG; Finogenova TV; Kondrashova MN (23 Oct 2006). "Evolution of glyoxylate cycle enzymes in Metazoa: evidence of multiple horizontal transfer events and pseudogene formation". Biol Direct. 1 (31): 31. PMC 1630690
 . PMID 17059607. doi:10.1186/1745-6150-1-31.
. PMID 17059607. doi:10.1186/1745-6150-1-31. - ↑ Kornberg, HL; Krebs HA (18 May 1957). "Synthesis of cell constituents from C2-units by a modified tricarboxylic acid cycle". Nature. 179 (4568): 988–991. PMID 13430766. doi:10.1038/179988a0.
- ↑ Idnurm, A; Howlett BJ (Oct 2002). "Isocitrate lyase is essential for pathogenicity of the fungus Leptosphaeria maculans to canola (Brassica napus)". Eukaryot Cell. 1 (5): 719–724. PMC 126752 . PMID 12455691. doi:10.1128/EC.1.5.719-724.2002.
- ↑ Lorenz, MC; Bender JA (Oct 2004). "Fink GR". Eukaryot Cell. 3 (5): 1076–1087. PMC 522606 . PMID 15470236. doi:10.1128/EC.3.5.1076-1087.2004.
- ↑ Srivastava, V; Jain A; Srivastava BS; Srivastava R (May 2008). "Selection of genes of Mycobacterium tuberculosis upregulated during residence in lungs of infected mice". Tuberculosis (Edinb). 88 (3): 171–177. PMID 18054522. doi:10.1016/j.tube.2007.10.002.
- ↑ Muñoz-Elías, EJ; McKinney JD (Jun 2005). "Mycobacterium tuberculosis isocitrate lyases 1 and 2 are jointly required for in vivo growth and virulence". Nat Med. 11 (6): 638–644. PMC 1464426 . PMID 15895072. doi:10.1038/nm1252.
Further reading
- McFadden BA; Howes WV (1963). "Crystallisation and some properties of isocitrate lyase from Pseudomonas indigofera". J. Biol. Chem. 238: 1737–1742.
- Shiio I, Shiio T; McFadden BA (1965). "Isocitrate lyase from Pseudomonas indigofera. I. Preparation, amino acid composition and molecular weight". Biochim. Biophys. Acta. 96: 114–22. PMID 14285253. doi:10.1016/0005-2787(65)90615-5.
- VICKERY HB (1962). "A suggested new nomenclature for the isomers of isocitric acid". J. Biol. Chem. 237: 1739–41. PMID 13925783.