Haplogroup R1a
| Haplogroup R1a | |
|---|---|
| Possible time of origin | 22,000 YBP [1] to 25,000[2] years ago |
| Possible place of origin | Eurasia (see text). |
| Ancestor | Haplogroup R1 |
| Descendants | Haplogroup R1a-Z282 (Europe), R1a-Z93 (Asia) |
| Defining mutations |
R1a: L62, L63, L120, M420, M449, M511, M513 R1a1a: M17, M198, M512, M514, M515, L168, L449, L457, L566 |
| Highest frequencies | See List of R1a frequency by population |
Haplogroup R1a, or haplogroup R-M420, is a human Y-chromosome DNA haplogroup which is distributed in a large region in Eurasia, extending from Scandinavia, Central Europe and southern Siberia to South Asia.[3][2]
While R1a originated ca. 22,000[1] to 25,000[2] years ago, its subclade M417 (R1a1a1) diversified ca. 5,800 years ago.[4] The distribution of M417-subclades R1-Z282 (including R1-Z280)[5] in Central and Eastern Europe and R1-Z93 in Asia[5][2] suggests that R1a1a diversified within the Eurasian Steppes or the Middle East and Caucasus region.[5] The place of origin of these subclades plays a role in the debate about the origins of Indo-Europeans.
The SNP mutation R-M420 was discovered after R-M17 (R1a1a), which resulted in a reorganization of the lineage in particular establishing a new paragroup (designated R-M420*) for the relatively rare lineages which are not in the R-SRY10831.2 (R1a1) branch leading to R-M17.
Origins
R1a origins
Tatiana et al. (2014) "rapid diversification process of K-M526 likely occurred in Southeast Asia, with subsequent westward expansions of the ancestors of haplogroups R and Q." [6]
The split of R1a (M420) is computed to ca. 22,000,[1] or 25,000[2] years ago, which is the time of the last glacial maximum. A large, 2014 study by Peter A Underhill et al., using 16,244 individuals from over 126 populations from across Eurasia, concluded that there was compelling evidence that "the initial episodes of haplogroup R1a diversification likely occurred in the vicinity of present-day Iran."[2]
Diversification of R1a1a1 (M417) and ancient migrations
_and_R1a1a_oldest_expansion_and_highest_frequency_(2014).jpg)
According to Underhill (2014), the downstream R1a-M417 subclade diversified into Z282 and Z93 circa 5,800 years ago.[4][note 1] The question of the origins of R1a1a is relevant to the ongoing debate concerning the urheimat of the proto-Indo-European people, and may also be relevant to the origins of the Indus Valley Civilisation. R1a shows a strong correlation with Indo-European languages of western/southern Asia and central/eastern Europe,[9][3] being most prevalent in Poland, Hungary, the Czech Republic, Slovakia, Belarus, Russia and Ukraine in Europe and in central Asia, Afghanistan, Pakistan and India in Asia. In Eastern Europe Z282 is prevalent, while in South Asia Z93 dominates. M45 is prevalent in Central Europe. The connection between Y-DNA R-M17 and the spread of Indo-European languages was first noted by T. Zerjal and colleagues in 1999.[10]
Steppe origins
Proposed steppe dispersal of R1a1a
Kivisild et al. (2003) have proposed either south or west Asia,[11][note 2] while Mirabal et al. (2009) see support for both south and central Asia.[9] Other studies suggest Ukrainian,[12] Central Asian[13] and West Asian origins for R1a1a.[14][3][15][5][2]
Ornella Semino et al. (2000) proposed Ukrainian origins, and a postglacial spread of the R1a1 gene during the Late Glacial Maximum, subsequently magnified by the expansion of the Kurgan culture into Europe and eastward.[16] Spencer Wells proposes central Asian origins, suggesting that the distribution and age of R1a1 points to an ancient migration corresponding to the spread by the Kurgan people in their expansion from the Eurasian steppe.[13] According to Pamjav et al. (2012), R1a1a diversified in the Eurasian Steppes or the Middle East and Caucasus region:
Inner and Central Asia is an overlap zone for the R1a1-Z280 and R1a1-Z93 lineages [which] implies that an early differentiation zone of R1a1-M198 conceivably occurred somewhere within the Eurasian Steppes or the Middle East and Caucasus region as they lie between South Asia and Central- and Eastern Europe."[5]
Three genetic studies in 2015 gave support to the Kurgan theory of Gimbutas regarding the Indo-European Urheimat. According to those studies, haplogroups R1b and R1a, now the most common in Europe (R1a is also common in South Asia) would have expanded from the Russian steppes, along with the Indo European languages; they also detected an autosomal component present in modern Europeans which was not present in Neolithic Europeans, which would have been introduced with paternal lineages R1b and R1a, as well as Indo European Languages.[17][18][19]
Source of R1a1a1 in Corded Ware culture

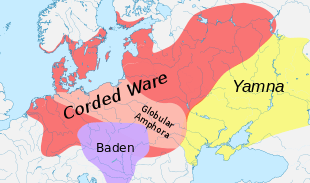

David Anthony considers the Yamna culture to be the Indo-European Urheimat.[20][21] According to Haak et al. (2015), a massive migration from the Yamna culture northwards took place ca. 2,500 BCE, accounting for 75% of the genetic ancestry of the Corded Ware culture, noting that R1a and R1b may have "spread into Europe from the East after 3,000 BCE".[22] Yet, all their seven Yamna samples belonged to the R1b-M269 subclade,[22] but no R1a1a has been found in their Yamna samples.[23] This raises the question where the R1a1a in the Corded Ware culture came from, if it was not from the Yamna culture.[24][25]
R1a may have migrated from the Anatolian-Iranian area via Central Asia to Eastern Europe, in concreto the Comb Ware culture (4,200 BCE – 2,000 BCE),[26] which was partly absorbed by the Corded Ware culture. R1a1 has been found in samples from the Narva culture,[26] which was part of the Comb Ware culture. Horvath rejects this possible migration route, given the dominance of haplogroup N1c in the Comb Ware culture, and that the Corded ware autosomal DNA is derived from the Yamna culture, and not from the Comb Ware culture.[26] In contrast, Semenov and Bulat do argue for such an origin of R1a1a in the Corded ware culture, noting that several publications point to the presence of R1a1 in the Comb Ware culture.[27][note 3]
Horvath proposes a migration of R1a from the Anatolian-Iranian area to the Pontic steppe via the Balkan.[24] Horvath notes that Haak et al. (2015) found that part of the Yamna ancestry derived from the Middle East, and that neolithic techniques probably arrived at the Yamna culture from the Balkans.[28][note 4] Horvath further notes that in the area of the Rossen culture (4,600–4,300 BC), which was situated on Germany and predates the Corded Ware culture, an old subclade of R1a, namely L664, can still be found.[30][note 5] From these facts Horvath speculates that R1a arrived in the Balkans via Anatolia, and from there spread first north-west to the Rossen culture, and then east from the Cucuteni culture to the Yamna and Afanasevo cultures, despite the absence of R1a from intermediate cultures between the Near East, Anatolia and the Balkans.[31][note 6][note 7]
Transcaucasia & West Asian origins and possible influence on Indus Valley Civilisation
Part of the South Asian genetic ancestry derives from west Eurasian populations, and some researchers have implied that Z93 may have come to India via Iran[33] and expanded there during the Indus Valley Civilisation.[2][34]
Mascarenhas et al. (2015) note that the roots of Z93 lie in West Asia, and propose that "Z93 and L342.2 expanded in a southeasterly direction from Transcaucasia into South Asia,"[33] noting that such an expansion is compatible with "the archeological records of eastward expansion of West Asian populations in the 4th millennium BCE culminating in the so-called Kura-Araxes migrations in the post-Uruk IV period."[33] Yet, Lazaridis noted that sample I1635 of Lazaridis et al. (2016), their Armenian Kura-Araxes sample, carried Y-haplogroup R1b1-M415(xM269)[note 8] (also called R1b1a1b-CTS3187).[35]
According to Underhill et al. (2014/2015) the diversification of Z93 and the "early urbanization within the Indus Valley [...] occurred at [5,600 years ago] and the geographic distribution of R1a-M780 (Figure 3d[note 9]) may reflect this."[2][note 10] Poznik et al. (2016) note that 'striking expansions' occurred within R1a-Z93 at ~4,500–4,000 years ago, which "predates by a few centuries the collapse of the Indus Valley Civilisation."[34]
Proposed South Asian origins
Kivisild et al. (2003) have proposed either South or West Asia,[11][note 2] while Mirabal et al. (2009) see support for both South and Central Asia.[9]
South Asian populations have the highest STR diversity within R1a1a,[37][38][9][3][1][39] and subsequent older TMRCA datings, and R1a1a is present among both higher (Brahmin) castes and lower castes, although the presence is substantially higher among Brahmin castes.[1][39] From these findings some researchers have concluded that R1a1a originated in South Asia,[38][1][note 11] excluding a substantial genetic influx from Indo-European migrants.[38][37][3]
However, this diversity, and the subsequent older TMRCA-datings, can also be explained by the historically high population numbers, which increases the likelihood of diversification and microsatellite variation.[41][42] According to Sengupta et al. (2006), "[R1a1 and R2] could have actually arrived in southern India from a southwestern Asian source region multiple times."[37][note 12] Silva et al. (2017) noted that R1a in South Asia most "likely spread from a single Central Asian source pool, there do seem to be at least three and probably more R1a founder clades within the Subcontinent, consistent with multiple waves of arrival."[42]
Phylogeny
The R1a family tree now has three major levels of branching, with the largest number of defined subclades within the dominant and best known branch, R1a1a (which will be found with various names; in particular, as "R1a1" in relatively recent but not the latest literature.)
Topology
The topology of R1a is as follows (codes [in brackets] non-isogg codes):[43][8][44][45][46] Tatiana et al. (2014) "rapid diversification process of K-M526 likely occurred in Southeast Asia, with subsequent westward expansions of the ancestors of haplogroups R and Q." [47]
- P P295/PF5866/S8 (also known as K2b2).
- R (R-M207)[43][8]
- R*
- R1 (R-M173)
- R1*[43]
- R1a (M420)[43] (Eastern Europe, Asia)[45]
- R1a*[8]
- R1a1[43] (M459/PF6235,[43] SRY1532.2/SRY10831.2[43])
- R1a1 (M459)[43][8]
- R1a1a (M17, M198)[43]
- R1a1a1 (M417, page7)[43]
- R1a1a1a (CTS7083/L664/S298)[43]
- R1a1a1b (S224/Z645, S441/Z647)[43]
- R1a1a1b1 (PF6217/S339/Z283)[43]
- R1a1a1b2 (F992/S202/Z93)[43] [R1a1a2*] (Z93, M746)(Asia)[48]
- [R1a1a1c] (M64.2, M87, M204)[46]
- [R1a1a1d] (P98)[46]
- [R1a1a1d2a][49]
- [R1a1a1e] (PK5)[46]
- R1a1a1 (M417, page7)[43]
- R1b (M343) (Western Europe)
- R2
Haplogroup R
|
R-M173 (R1)
R1a is distinguished by several unique markers, including the M420 mutation. It is a subclade of Haplogroup R-M173 (previously called R1). R1a has the sister-subclades Haplogroup R1b-M343, and the paragroup R-M173*.
R-M420 (R1a)
R-M420, defined by the mutation M420, has two branches: R-SRY1532.2, defined by the mutation SRY1532.2, which makes up the vast majority; and R-M420*, the paragroup, defined as M420 positive but SRY1532.2 negative. (In the 2002 scheme, this SRY1532.2 negative minority was one part of the relatively rare group classified as the paragroup R1*.) Mutations understood to be equivalent to M420 include M449, M511, M513, L62, and L63.[3][50]
Only isolated samples of the new paragroup R-M420* were found by Underhill 2009, mostly in the Middle East and Caucasus: 1/121 Omanis, 2/150 Iranians, 1/164 in the United Arab Emirates, and 3/612 in Turkey. Testing of 7224 more males in 73 other Eurasian populations showed no sign of this category.[3]
R-SRY1532.2 (R1a1)
R1a1 is defined by SRY1532.2 or SRY10831.2), understood to always include SRY10831.2, M448, L122, M459, and M516.[3][51]) This family of lineages is dominated by M17 and M198. In contrast, paragroup R-SRY1532.2* lacks either the M17 or M198 markers.
The R-SRY1532.2* paragroup is apparently less rare than R1*, but still relatively unusual, though it has been tested in more than one survey. Underhill et all. (2009) reported 1/51 in Norway, 3/305 in Sweden, 1/57 Greek Macedonians, 1/150 Iranians, 2/734 ethnic Armenians, and 1/141 Kabardians.[3] Sahoo et al. (2006) reported R-SRY1532.2* for 1/15 Himachal Pradesh Rajput samples.[38]
R-M17/M198 (R1a1a)
The following SNPs are associated with R1a1a:
| SNP | Mutation | Y-position (NCBI36) | Y-position (GRCh37) | RefSNP ID |
|---|---|---|---|---|
| M17 | INS G | 20192556 | 21733168 | rs3908 |
| M198 | C->T | 13540146 | 15030752 | rs2020857 |
| M512 | C->T | 14824547 | 16315153 | rs17222146 |
| M514 | C->T | 17884688 | 19375294 | rs17315926 |
| M515 | T->A | 12564623 | 14054623 | rs17221601 |
| L168 | A->G | 14711571 | 16202177 | - |
| L449 | C->T | 21376144 | 22966756 | - |
| L457 | G->A | 14946266 | 16436872 | rs113195541 |
| L566 | C->T | - | - | - |
R-M17 (R1a1a1)
R1a1a1 (RM-417) is the most widely found subclade, in two variations which are found respectively in Europe (R1a1a1b1 (R-Z282) ([R1a1a1a*] (R-Z282) (Underhill 2014/2015)[45]) and Central and South Asia (R1a1a1b2 (R-Z93) ([R1a1a2*] (R-Z93) Underhill 2014/2015)[45]).
R-Z282 (R1a1a1b1a) (Eastern Europe)
This large subclade appears to encompass most of the R1a1a found in Europe.[52]
- R1a1a1b1a [R1a1a1a* (Underhill (2014))] (R-Z282*) occurs in northern Ukraine, Belarus, and Russia at a frequency of ~20%. (Underhill et al. 2014)
- R1a1a1b1a3 [R1a1a1a1 (Underhill (2014))] (R-Z284) occurs in Northwest Europe and peaks at ~20% in Norway. (Underhill et al. 2014)
- R1a1a1c (M64.2, M87, M204) is apparently rare: it was found in 1 of 117 males typed in southern Iran.[14]
R-M458 (R1a1a1b1a1)
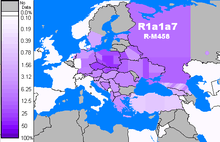
R-M458 is a mainly Slavic SNP, characterized by its own mutation, and was first called cluster N. Underhill et al. (2009) found it to be present in modern European populations roughly between the Rhine catchment and the Ural Mountains and traced it to "a founder effect that [...] falls into the early Holocene period, 7.9±2.6 KYA."[53] M458 was found in one skeleton from a 14th-century grave field in Usedom, Mecklenburg-Vorpommern, Germany.[54] The paper by Underhill et al. (2009) also reports a surprisingly high frequency of M458 in some Northern Caucasian populations (for example 27.5% among Karachays and 23.5% among Balkars, 7.8% among Karanogays and 3.4% among Abazas).
R-L260 (R1a1a1b1a1a) (Gwozdz's cluster P)
R1a1a1b1a1a (R-L260), commonly referred to as West Slavic or Polish, is a subclade of the larger parent group R-M458, and was first identified as an STR cluster by Pawlowski 2002 and then by Gwozdz 2009. Thus, R-L260 was what Gwozdz 2009 called cluster "P." In 2010 it was verified to be a haplogroup identified by its own mutation (SNP).[55] It apparently accounts for about 8% of Polish men, making it the most common subclade in Poland. Outside of Poland it is less common (Pawlowski 2002). In addition to Poland, it is mainly found in the Czech Republic and Slovakia, and is considered "clearly West Slavic."[56] The founding ancestor of R-L260 is estimated to have lived between 2000 and 3000 years ago, i.e. during the Iron Age, with significant population expansion less than 1,500 years ago.[57]
R-M334
R-M334 ([R1a1a1g1],[46] a subclade of [R1a1a1g] (M458)[46] c.q. R1a1a1b1a1 (M458)[43]) was found by Underhill et al. (2009) only in one Estonian man and may define a very recently founded and small clade.[3]
R1a1a1b1a2 (S466/Z280, S204/Z91)
R1a1a1b1a2b3* (Gwozdz's Cluster K)
R1a1a1b1a2b3* (M417+, Z645+, Z283+, Z282+, Z280+, CTS1211+, CTS3402, Y33+, CTS3318+, Y2613+) (Gwozdz's Cluster K)[44] is a STR based group that is R-M17(xM458). This cluster is common in Poland but not exclusive to Poland.[57]
R1a1a1b1a2b3a (R-L365)
R1a1a1b1a2b3a (R-L365)[43] was early called Cluster G.
R1a1a1b2 (R-Z93) (Asia)
This large subclade appears to encompass most of the R1a1a found in Asia.[52]
- R1a1a1b2 [R1a1a2* (Underhill (2014))] (R-Z93) is most common (>30%) in the South Siberian Altai region of Russia, cropping up in Kyrgyzstan (6%) and in all Iranian populations (1–8%).[58]
- R1a1a1b2a* (R-Z2125): This subgroup occurs at highest frequencies in Kyrgyzstan and in Afghan Pashtuns (>40%). At a frequency of >10% it is also observed in other Afghan ethnic groups and in some populations in the Caucasus and Iran.[58]
| Region | People | N | R-M17 | R-M434 | ||
| Number | Freq. (%) | Number | Freq. (%) | |||
| Pakistan | Baloch | 60 | 9 | 15% | 5 | 8% |
| Pakistan | Makrani | 60 | 15 | 25% | 4 | 7% |
| Middle East | Oman | 121 | 11 | 9% | 3 | 2.5% |
| Pakistan | Sindhi | 134 | 65 | 49% | 2 | 1% |
| Table only shows positive sets from N = 3667 derived from 60 Eurasian populations sample.[3] | ||||||
- R-M434 is a subclade of Z2125. It was detected in 14 people (out of 3667 people tested) all in a restricted geographical range from Pakistan to Oman. This likely reflects a recent mutation event in Pakistan (Underhill 2009).
- R1a1b2a1* (R-M560 is very rare and was only observed in four samples: two Burushaski speakers (north Pakistan), one Hazara (Afghanistan), and one Iranian Azerbaijani.[58]
- R1a1b2a2* (R-M780) occurs at high frequency in South Asia: India, Pakistan, Afghanistan, and the Himalayas. The group also occurs at >3% in some Iranian populations and is present at >30% in Roma from Croatia and Hungary.[58]
Geographic distribution of R1a1a



Historical
Haplogroup R1a has been found in ancient fossils associated with the Corded Ware culture[59][60] and Urnfield culture;[61] as well as the burial of the remains of the Sintashta culture,[62] Andronovo culture,[63] the Pazyryk culture,[64] Tagar culture[63] and Tashtyk culture,[63] the inhabitants of ancient Tanais,[65] in the Tarim mummies,[66] the aristocracy Xiongnu,[67] in two ancient Khazar fossils. The skeletal remains of a father and his two sons, from an archaeological site discovered in 2005 near Eulau (in Saxony-Anhalt, Germany) and dated to about 2600 BCE, tested positive for the Y-SNP marker SRY10831.2. The Ysearch number for the Eulau remains is 2C46S. The ancestral clade was thus present in Europe at least 4600 years ago, in association with one site of the widespread Corded Ware culture.[68]
Europe
In Europe, the R1a1 sub-clade is found at highest levels among peoples of Eastern European descent, with 50 to 65% among Sorbs, Poles, Russians and Ukrainians.[69][70][12] In the Baltic countries R1a1a frequencies decrease from Lithuania (45%) to Estonia (around 30%).[71] Levels in Hungarians have been noted between 20 and 60%.[72][73][12][74]
There is a significant presence in peoples of Scandinavian descent, with highest levels in Norway and Iceland, where between 20 and 30% of men are in R1a1a.[75][76] Vikings and Normans may have also carried the R1a1a lineage westward; accounting for at least part of the small presence in the British Isles.[77][78] In East Germany, where Haplogroup R1a1a reaches a peak frequency in Rostock at a percentage of 31.3%, it averages between 20 and 30%.[79]
Haplogroup R1a1a was found at elevated levels among a sample of the Israeli population who self-designated themselves as Levites and Ashkenazi Jews (Levites comprise approximately 4% of Jews). Behar reported R1a1a to be the dominant haplogroup in Ashkenazi Levites (52%), although rare in Ashkenazi Cohanim (1.3%).[70]
In Southern Europe R1a1a is not common, but significant levels have been found in pockets, such as in the Pas Valley in Northern Spain, areas of Venice, and Calabria in Italy.[80] The Balkans shows lower frequencies, and significant variation between areas, for example more than 30% in Slovenia, Croatia and Greek Macedonia, but less than 10% in Albania, Kosovo and parts of Greece.[81][73][12]
R1a is virtually composed only of the Z284 subclade in Scandinavia, which is only found in single sample of a Slovenian in Eastern Europe, where the main subclade is Z282(Z280 and M458) and there is a negligible representation of Z93 in each region other than Turkey.[45] The West Slavs and Hungarians are characterized by a high frequency of the subclade M458 and a low Z92, a subclade of Z280. Hundreds of samples of each Slovenians, and Czechs lack the Z92 subclade of Z280, while Poles, Slovaks, Croats and Hungarians only show a very low frequency of Z92.[45] The Balts, East Slavs, Serbs, Macedonians, Bulgarians and Romanians demonstrate a ratio Z280>M458 and a high, up to a prevailing share of Z92.[45] Balts and East Slavs have the same subclades and similar frequencies in a more detailed phylogeny of the subclades.[82][83] The Russian geneticist Oleg Balanovsky speculated that there is a predominance of the assimilated pre-Slavic substrate in the genetics of East and West Slavic populations, according to him the common genetic structure which contrasts East Slavs and Balts frоm other populations may suggest the explanation that the pre-Slavic substrate of the East Slavs consisted most significantly of Baltic-speakers, which at one point predated the Slavs in the cultures of the Eurasian steppe according to archaeological and toponymic references.[84]
Asia
Central Asia
In Afghanistan, R1a1a is found at 51% among the Pashtuns who are the largest ethnic group in Afghanistan, 50% among the Kyrgyz, and 30% among the Tajiks. It is less frequent among the Hazaras (7%) and the Uzbeks (18%).[85]
South Asia
In South Asia, R1a1a has often been observed with high frequency in a number of demographic groups.[38][37]
In India, high frequencies of this haplogroup is observed in West Bengal Brahmins (72%)[37] to the east, Konkanastha Brahmins (48%)[37] to the west, Khatris (67%)[3] in the north and Iyengar Brahmins (31%)[37] in the south. It has also been found in several South Indian Dravidian-speaking Adivasis including the Chenchu (26%) and the Valmikis of Andhra Pradesh and the Kallar of Tamil Nadu suggesting that R1a1a is widespread in Tribal Southern Indians.[11]
Besides these, studies show high percentages in regionally diverse groups such as Manipuris (50%)[3] to the extreme North East and among Punjabis (47%)[11] to the extreme North West.
In Pakistan it is found at 71% among the Mohanna tribe in Sindh province to the south and 46% among the Baltis of Gilgit-Baltistan to the north.[3] Among the Sinhalese of Sri Lanka, 23% were found to be R1a1a (R-SRY1532) positive. [86] Hindus of Chitwan District in the Terai region Nepal show it at 69%.[87]
East Asia
The frequency of R1a1a is comparatively low among some Turkic-speaking groups including Turks, Azeris, Kazakhs, and Yakuts, yet levels are higher (19 to 28%) in certain Turkic or Mongolic-speaking groups of Northwestern China, such as the Bonan, Dongxiang, Salar, and Uyghurs.[13][88][89]
In Eastern Siberia, R1a1a is found among certain indigenous ethnic groups including Kamchatkans and Chukotkans, and peaking in Itel'man at 22%.[90]
West Asia
R1a1a has been found in various forms, in most parts of Western Asia, in widely varying concentrations, from almost no presence in areas such as Jordan, to much higher levels in parts of Kuwait, Turkey and Iran. The Shimar (Shammar) Bedouin tribe in Kuwait show the highest frequency in the Middle East at 43%.[91][92][93])
Wells 2001, noted that in the western part of the country, Iranians show low R1a1a levels, while males of eastern parts of Iran carried up to 35% R1a1a. Nasidze 2004 found R1a1a in approximately 20% of Iranian males from the cities of Tehran and Isfahan. Regueiro 2006 in a study of Iran, noted much higher frequencies in the south than the north.
A newer Study has found 20.3% R-M17* among Kurdish samples which were taken in the Kurdistan Province in western Iran, 9.7% among Mazandaranis in North Iran in the province of Mazandaran, 9.4% among Gilaks in province of Gilan, 12.8% among Persian and 17.6% among Zoroastrians in Yazd, 18.2% among Persians in Isfahan, 20.3% among Persians in Khorasan, 16.7% Afro-Iranians, 18.4% Qeshmi "Gheshmi", 21.4% among Persian Speaking Bandari people in Hormozgan and 25% among the Baloch people in Sistan and Baluchestan Province.[94]
Further to the north of these Middle Eastern regions on the other hand, R1a1a levels start to increase in the Caucasus, once again in an uneven way. Several populations studied have shown no sign of R1a1a, while highest levels so far discovered in the region appears to belong to speakers of the Karachay-Balkar language among whom about one quarter of men tested so far are in haplogroup R1a1a.[3]
Popular science
Bryan Sykes in his book Blood of the Isles gives imaginative names to the founders or "clan patriarchs" of major British Y haplogroups, much as he did for mitochondrial haplogroups in his work The Seven Daughters of Eve. He named R1a1a in Europe the "clan" of a "patriarch" Sigurd, reflecting the theory that R1a1a in the British Isles has Norse origins.
In art
Artem Lukichev created an animation based on the Bashkir epic about the Ural, which outlined the history of the clusters of haplogroup R1: R1a and R1b.[95]
Historic naming of "R1a"
The historic naming system commonly used for R1a was inconsistent in different published sources, because it changed often; this requires some explanation.
In 2002, the Y Chromosome Consortium (YCC) proposed a new naming system for haplogroups (YCC 2002), which has now become standard. In this system, names with the format "R1" and "R1a" are "phylogenetic" names, aimed at marking positions in a family tree. Names of SNP mutations can also be used to name clades or haplogroups. For example, as M173 is currently the defining mutation of R1, R1 is also R-M173, a "mutational" clade name. When a new branching in a tree is discovered, some phylogenetic names will change, but by definition all mutational names will remain the same.
The widely occurring haplogroup defined by mutation M17 was known by various names, such as "Eu19", as used in (Semino 2000) in the older naming systems. The 2002 YCC proposal assigned the name R1a to the haplogroup defined by mutation SRY1532.2. This included Eu19 (i.e. R-M17) as a subclade, so Eu19 was named R1a1. Note, SRY1532.2 is also known as SRY10831.2 The discovery of M420 in 2009 has caused a reassignment of these phylogenetic names.(Underhill 2009 and ISOGG 2012) R1a is now defined by the M420 mutation: in this updated tree, the subclade defined by SRY1532.2 has moved from R1a to R1a1, and Eu19 (R-M17) from R1a1 to R1a1a.
More recent updates recorded at the ISOGG reference webpage involve branches of R-M17, including one major branch, R-M417.
| 2002 Scheme proposed in (YCC 2002) | 2009 Scheme as per (2009) | Latest ISOGG tree as per January 2011 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
|
|
See also
Y-DNA R-M207 subclades
Y-DNA backbone tree
| Phylogenetic tree of human Y-chromosome DNA haplogroups [χ 1][χ 2] | |||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| "Y-chromosomal Adam" | |||||||||||||||||||||||||||||||||||||||||||||||||
| A00 | A0-T [χ 3] | ||||||||||||||||||||||||||||||||||||||||||||||||
| A0 | A1 [χ 4] | ||||||||||||||||||||||||||||||||||||||||||||||||
| A1a | A1b | ||||||||||||||||||||||||||||||||||||||||||||||||
| A1b1 | BT | ||||||||||||||||||||||||||||||||||||||||||||||||
| B | CT | ||||||||||||||||||||||||||||||||||||||||||||||||
| DE | CF | ||||||||||||||||||||||||||||||||||||||||||||||||
| D | E | C | F | ||||||||||||||||||||||||||||||||||||||||||||||
| F1 | F2 | F3 | GHIJK | ||||||||||||||||||||||||||||||||||||||||||||||
| G | HIJK | ||||||||||||||||||||||||||||||||||||||||||||||||
| IJK | H | ||||||||||||||||||||||||||||||||||||||||||||||||
| IJ | K | ||||||||||||||||||||||||||||||||||||||||||||||||
| I | J | LT [χ 5] | K2 | ||||||||||||||||||||||||||||||||||||||||||||||
| L | T [χ 6] | K2a [χ 7] | K2b [χ 8] | K2c | K2d | K2e [χ 9] | |||||||||||||||||||||||||||||||||||||||||||
| K2a1 | K2b1 [χ 10] | P [χ 11] | |||||||||||||||||||||||||||||||||||||||||||||||
| NO | S [χ 12] | M [χ 13] | P1 | P2 | |||||||||||||||||||||||||||||||||||||||||||||
| NO1 | Q | R | |||||||||||||||||||||||||||||||||||||||||||||||
| N | O | ||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||||||||||||||||||||
Notes
- ↑ According to Family Tree, they diversified ca. 5,000 years ago.[8]
- 1 2 Kivisild et al. (2003): "Haplogroup R1a, previously associated with the putative Indo-Aryan invasion, was found at its highest frequency in Punjab but also at a relatively high frequency (26%) in the Chenchu tribe. This finding, together with the higher R1a-associated short tandem repeat diversity in India and Iran compared with Europe and central Asia, suggests that southern and western Asia might be the source of this haplogroup."[11]
- ↑ Semenov and Bulat refer to the following publications:
5. Haak W. et al. Massive migration from the steppe is a source for Indo-European languages in Europe. doi:10.1101/013433.
6. Mathieson I et al. Eight thousand years of natural selection in Europe. doi:10.1101/016477
8. Chekunova Е.М., Yartseva N.V., Chekunov М.К., Мazurkevich А.N. The First Results of the Genotyping of the Aboriginals and Human Bone Remains of the Archeological Memorials of the Upper Podvin’e. // Archeology of the lake settlements of IV—II Thousands BC: The chronology of cultures and natural environment and climatic rhythms. Proceedings of the International Conference, Devoted to the 50-year Research of the Pile Settlements on the North-West of Russia. St. Petersburg, 13–15 November 2014.
9. Eppie R. Jones et al. Upper Palaeolithic genomes reveal deep roots of modern Eurasians. Nature Communications. doi:10.1038/ncomms9912 PMID 26567969 - ↑ Yet, Haak et al. also explicitly state: "...a type of Near Eastern ancestry different from that which was introduced by early farmers."[29]
- ↑ According to Family Tree DNA, L664 formed 4,700 ybp, that is, 2,700 BCE.[8]
- ↑ Asko Parpola (2015) proposes the Cucuteni-Trypolye culture as the carrier of late Proto-Indo-European. He notes that the Cucuteni-Trypolye culture may have been the birthplace of wheeled vehicles, giving the words related to these vehicles. Parpola further notes that the Cucuteni-Trypolye culture was taken over by PIE speakers at circa 4,000 BCE, and expanded to the Pontic steppe ca. 3,400 BCE, eventually giving rise to the Yamna culture.[32]
- ↑ See Eupedia.com for some critical comments on Horvath (2016).
- ↑ Lazaridis, Twitter, 18 juni 2016: "I1635 (Armenia_EBA) is R1b1-M415(xM269). We'll be sure to include in the revision. Thanks to the person who noticed! #ILovePreprints."
See also Eurogenes Blog, Big deal of 2016: the territory of present-day Iran cannot be the Indo-European homeland, for a discussion of the same topic. - ↑ See map for M780 distribution at Dieneke's Anthropology Blog, Major new article on the deep origins of Y-haplogroup R1a (Underhill et al. 2014)[36]
- ↑ According to Family Tree DNA, M780 formed 4700 ybp.[8] This dating coincides with the eastward movement between 2800 and 2600 BCE of the Yamna culture into the region of the Poltavka culture, a predecessor of the Sintashta culture, from which the Indo-Iranians originated. M780 is concentrated in the Ganges Vally, the locus of the classic Vedic society.
- ↑ Qutes:
- Sahoo et al. (2006): "... one should expect to observe dramatically lower genetic variation among Indian Rla lineages. In fact, the opposite is true: the STR haplotype diversity on the background of R1a in Central Asia (and also in Eastern Europe) has already been shown to be lower than that in India (6). Rather, the high incidence of R1* and Rla throughout Central Asian European populations (without R2 and R* in most cases) is more parsimoniously explained by gene flow in the opposite direction, possibly with an early founder effect in South or West Asia.[40]
- Sharma et al. (2009): "A peculiar observation of the highest frequency (up to 72.22%) of Y-haplogroup R1a1* in Brahmins hinted at its presence as a founder lineage for this caste group. Further, observation of R1a1* in different tribal population groups, existence of Y-haplogroup R1a* in ancestors and extended phylogenetic analyses of the pooled dataset of 530 Indians, 224 Pakistanis and 276 Central Asians and Eurasians bearing the R1a1* haplogroup supported the autochthonous origin of R1a1 lineage in India and a tribal link to Indian Brahmins. However, it is important to discover novel Y-chromosomal binary marker(s) for a higher resolution of R1a1* and confirm the present conclusions."[1]
- ↑ Sengupta et al. (2006): "The widespread geographic distribution of HG R1a1-M17 across Eurasia and the current absence of informative subdivisions defined by binary markers leave uncertain the geographic origin of HG R1a1-M17. However, the contour map of R1a1-M17 variance shows the highest variance in the northwestern region of India [...] The question remains of how distinctive is the history of L1 relative to some or all of R1a1 and R2 representatives. This uncertainty neutralizes previous conclusions that the intrusion of HGs R1a1 and R2 from the northwest in Dravidian-speaking southern tribes is attributable to a single recent event. [R1a1 and R2] could have actually arrived in southern India from a southwestern Asian source region multiple times, with some episodes considerably earlier than others. Considerable archeological evidence exists regarding the presence of Mesolithic peoples in India (Kennedy 2000), some of whom could have entered the subcontinent from the northwest during the late Pleistocene epoch. The high variance of R1a1 in India (table 12), the spatial frequency distribution of R1a1 microsatellite variance clines (fig. 4), and expansion time (table 11) support this view."[37]
References
- 1 2 3 4 5 6 7 Sharma 2009.
- 1 2 3 4 5 6 7 8 9 Underhill 2014.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 Underhill 2009.
- 1 2 Underhill 2014, p. 130.
- 1 2 3 4 5 Pamjav 2012.
- ↑ Karafet, Tatiana; Mendez, Fernando; Sudoyo, Herawati (2014). "Improved phylogenetic resolution and rapid diversification of Y-chromosome haplogroup K-M526 in Southeast Asia". Nature. 23: 369–373. PMC 4326703
 . PMID 24896152. doi:10.1038/ejhg.2014.106.
. PMID 24896152. doi:10.1038/ejhg.2014.106. - ↑ Underhill, PA; Myres, NM; Rootsi, S; Metspalu, M; Zhivotovsky, LA; King, RJ; Lin, AA; Chow, CE; Semino, O; Battaglia, V; Kutuev, I; Järve, M; Chaubey, G; Ayub, Q; Mohyuddin, A; Mehdi, SQ; Sengupta, S; Rogaev, EI; Khusnutdinova, EK; Pshenichnov, A; Balanovsky, O; Balanovska, E; Jeran, N; Augustin, DH; Baldovic, M; Herrera, RJ; Thangaraj, K; Singh, V; Singh, L; Majumder, P; Rudan, P; Primorac, D; Villems, R; Kivisild, T (2010). "Separating the post-Glacial coancestry of European and Asian Y chromosomes within haplogroup R1a". Eur. J. Hum. Genet. 18: 479–84. PMC 2987245 . PMID 19888303. doi:10.1038/ejhg.2009.194.
- 1 2 3 4 5 6 7 8 9 10 yfull.com, R1a tree
- 1 2 3 4 Mirabal 2009.
- ↑ T. Zerjal et al, The use of Y-chromosomal DNA variation to investigate population history: recent male spread in Asia and Europe, in S.S. Papiha, R. Deka and R. Chakraborty (eds.), Genomic diversity: applications in human population genetics (1999), pp. 91–101.
- 1 2 3 4 5 Kivisild 2003.
- 1 2 3 4 Semino 2000.
- 1 2 3 Wells 2001.
- 1 2 Regueiro 2006.
- ↑ Zhao 2009.
- ↑ Ornella Semino, Giuseppe Passarino, Peter J. Oefner, Alice A. Lin, Svetlana Arbuzova, Lars E. Beckman, Giovanna De Benedictis, Paolo Francalacci, Anastasia Kouvatsi, Svetlana Limborska, Mladen Marciki, Anna Mika, Barbara Mika, Dragan Primorac, A. Silvana Santachiara-Benerecetti, L. Luca Cavalli-Sforza, Peter A. Underhill, The Genetic Legacy of Paleolithic Homo sapiens sapiens in Extant Europeans: A Y Chromosome Perspective, Science, vol. 290 (10 November 2000), pp. 1155–1159.
- ↑ Massive migration from the steppe is a source for Indo-European languages in Europe, Haak et al, 2015
- ↑ Population genomics of Bronze Age Eurasia, Allentoft et al, 2015
- ↑ Eight thousand years of natural selection in Europe, Mathieson et al, 2015
- ↑ Anthony 2007.
- ↑ Anthony & Ringe 2015.
- 1 2 Haak 2015, p. 5.
- ↑ Horvath 2016, p. 199.
- 1 2 Horvath 2016.
- ↑ Semenov & Bulat 2016.
- 1 2 3 Horvath 2016, p. 201.
- ↑ Semenov & Bulat 2016, p. 41.
- ↑ Horvath 2016, p. 2015.
- ↑ Haak 2015, p. 4.
- ↑ Horvath.
- ↑ Horvath 2016, p. 197.
- ↑ Parpola 2015, p. 43-47.
- 1 2 3 Mascarenhas 2015, p. 9.
- 1 2 Pozink 2016, p. 5.
- ↑ Arame's English blog, Y DNA from ancient Near East
- ↑ Major new article on the deep origins of Y-haplogroup R1a (Underhill et al. 2014)
- 1 2 3 4 5 6 7 8 Sengupta 2006.
- 1 2 3 4 5 Sahoo 2006.
- 1 2 Thangaraj 2010.
- ↑ Sahoo 2006, p. 845-846.
- ↑ Joseph 2017.
- 1 2 Silva 2017.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 ISOGG, Y-DNA Haplogroup R and its Subclades – 2016
- 1 2 3 familytreedna.com, R1a-project
- 1 2 3 4 5 6 7 8 Underhill 2015.
- 1 2 3 4 5 6 7 8 9 10 11 snpedia, Haplogroup R (Y-DNA)
- ↑ Karafet, Tatiana; Mendez, Fernando; Sudoyo, Herawati (2014). "Improved phylogenetic resolution and rapid diversification of Y-chromosome haplogroup K-M526 in Southeast Asia". Nature. 23: 369–373. PMC 4326703 . PMID 24896152. doi:10.1038/ejhg.2014.106.
- 1 2 3 4 5 6 7 8 9 10 11 12 13 Underhill 2015, p. 125.
- ↑ eurogenes.blogspot, R1a in Yamnaya
- ↑ ISOGG 2012.
- ↑ Krahn 2012.
- 1 2 (Pamjav 2012).
- ↑ Underhill 2010.
- ↑ J. Freder, Die mittelalterlichen Skelette von Usedom [The mediaeval skeletons of Usedom], Berlin 2010, p. 86 (Dissertation Free University Berlin 2010).
- ↑ Peter Gwozdw. M458, L260, CTS11962
- ↑ Haplogroup R1a (Y-DNA)
- 1 2 Gwozdz 2009.
- 1 2 3 4 Underhill et al. 2014
- ↑ Haak, Wolfgang; Brandt, Guido; Jong, Hylke N. de; Meyer, Christian; Ganslmeier, Robert; Heyd, Volker; Hawkesworth, Chris; Pike, Alistair W. G.; Meller, Harald; Alt, Kurt W. (25 November 2008). "Ancient DNA, Strontium isotopes, and osteological analyses shed light on social and kinship organization of the Later Stone Age". PNAS. 105 (47): 18226–18231. PMC 2587582 . PMID 19015520. doi:10.1073/pnas.0807592105. Retrieved 15 June 2016 – via www.pnas.org.
- ↑ Brandit, G (2013). "Ancient DNA Reveals Key Stages in the Formation of Central European Mitochondrial Genetic Diversity". Science. 342 (6155): 257–261. PMC 4039305 . PMID 24115443. doi:10.1126/science.1241844.
- ↑ Schweitzer, D. (23 March 2008). "Lichtenstein Cave Data Analysis" (PDF). dirkschweitzer.net. Archived from the original (PDF) on 14 August 2011.
- ↑ Allentoft 2015.
- 1 2 3 Keyser, Christine; Bouakaze, Caroline; Crubézy, Eric; Nikolaev, Valery G.; Montagnon, Daniel; Reis, Tatiana; Ludes, Bertrand (2009). "Ancient DNA provides new insights into the history of south Siberian Kurgan people". Human Genetics. 126 (3): 395–410. ISSN 0340-6717. PMID 19449030. doi:10.1007/s00439-009-0683-0.
- ↑ Ricaut, F.; et al. (2004). "Genetic Analysis of a Scytho-Siberian Skeleton and Its Implications for Ancient Central Asian Migrations". Human Biology. 76: 1.
- ↑ Корниенко И. В., Водолажский Д. И. Использование нерекомбинантных маркеров Y-хромосомы в исследованиях древних популяций (на примере поселения Танаис)//Материалы Донских антропологических чтений. Ростов-на-Дону, Ростовский научно-исследовательский онкологический институт, Ростов-на-Дону, 2013.
- ↑ Chunxiang, Li; et al. (2010). "Evidence that a West-East admixed population lived in the Tarim Basin as early as the early Bronze Age" (PDF). BMC Biology. 8 (1): 15. ISSN 1741-7007. PMC 2838831 . PMID 20163704. doi:10.1186/1741-7007-8-15. Archived from the original (PDF) on 27 April 2011.
- ↑ Kim, Kijeong; Brenner, Charles H.; Mair, Victor H.; Lee, Kwang-Ho; Kim, Jae-Hyun; Gelegdorj, Eregzen; Batbold, Natsag; Song, Yi-Chung; Yun, Hyeung-Won; Chang, Eun-Jeong; Lkhagvasuren, Gavaachimed; Bazarragchaa, Munkhtsetseg; Park, Ae-Ja; Lim, Inja; Hong, Yun-Pyo; Kim, Wonyong; Chung, Sang-In; Kim, Dae-Jin; Chung, Yoon-Hee; Kim, Sung-Su; Lee, Won-Bok; Kim, Kyung-Yong (2010). "A western Eurasian male is found in 2000-year-old elite Xiongnu cemetery in Northeast Mongolia". American Journal of Physical Anthropology. 142 (3): 429–440. ISSN 0002-9483. PMID 20091844. doi:10.1002/ajpa.21242.
- ↑ Haak 2008.
- ↑ Balanovsky 2008.
- 1 2 Behar 2003.
- ↑ Kasperaviciūte 2005.
- ↑ Battaglia 2008.
- 1 2 Rosser 2000.
- ↑ Tambets 2004.
- ↑ Bowden 2008.
- ↑ Dupuy 2005.
- ↑ Passarino 2002.
- ↑ Capelli 2003.
- ↑ Kayser 2005.
- ↑ Scozzari 2001.
- ↑ Pericić 2005.
- ↑ http://pereformat.ru/wp-content/uploads/2015/02/russian-plain-01.jpg. Missing or empty
|title=(help) - ↑ http://www.rodstvo.ru/forum/index.php?act=attach&type=post&id=1299. Missing or empty
|title=(help) - ↑ П, Балановский О. "Генофонд Европы" (in Russian). KMK Scientific Press.
Прежде всего, это преобладание в славянских популяциях дославянского субстрата — двух ассимилированных ими генетических компонентов – восточноевропейского для западных и восточных славян и южноевропейского для южных славян...Можно с осторожностью предположить, что ассимилированный субстратмог быть представлен по преимуществу балтоязычными популяциями. Действительно, археологические данные указыва ют на очень широкое распространение балтских групп перед началом расселения славян. Балтскийсубстрату славян (правда, наряду с финно-угорским) выявляли и антропологи. Полученные нами генетические данные — и на графиках генетических взаимоотношений, и по доле общих фрагментов генома — указывают, что современные балтские народы являются ближайшими генетически ми соседями восточных славян. При этом балты являются и лингвистически ближайшими род ственниками славян. И можно полагать, что к моменту ассимиляции их генофонд не так сильно отличался от генофонда начавших свое широкое расселение славян. Поэтому если предположить,что расселяющиеся на восток славяне ассимилировали по преимуществу балтов, это может объяснить и сходство современных славянских и балтских народов друг с другом, и их отличия от окружающих их не балто-славянских групп Европы...В работе высказывается осторожное предположение, что ассимилированный субстрат мог быть представлен по преимуществу балтоязычными популяциями. Действительно, археологические данные указывают на очень широкое распространение балтских групп перед началом расселения славян. Балтский субстрат у славян (правда, наряду с финно-угорским) выявляли и антропологи. Полученные в этой работе генетические данные — и на графиках генетических взаимоотношений, и по доле общих фрагментов генома — указывают, что современные балтские народы являются ближайшими генетическими соседями восточных славян.
- ↑ Haber 2012.
- ↑ "The Genetics of Language and Farming Spread in India" (PDF).
- ↑ Fornarino 2009.
- ↑ Wang 2003.
- ↑ Zhou 2007.
- ↑ Lell 2002.
- ↑ Mohammad 2009.
- ↑ Nasidze 2004.
- ↑ Nasidze 2005.
- ↑ Grugni 2012.
- ↑ Lukichev, Artem (5 August 2014). "About R1a and R1b from Ural epic story". Retrieved 15 June 2016 – via YouTube.
{kind=link}
Sources
- Allentoft; et al. (2015), "Population genomics of Bronze Age Eurasia", Nature, 522: 167–172, doi:10.1038/nature14507
- Anthony, David W. (2007), The Horse The Wheel And Language. How Bronze-Age Riders From the Eurasian Steppes Shaped The Modern World, Princeton University Press
- Anthony, David; Ringe, Don (2015), "The Indo-European Homeland from Linguistic and Archaeological Perspectives", Annual Review of Linguistics, 1: 199–219, doi:10.1146/annurev-linguist-030514-124812
- Horvath, Csaba Barnabas (2016), "R1a subclades and Bronze Age migrations on the Eurasian steppes", European Scientific Journal, 12 (30)
- "Y-DNA Haplogroup R and its Subclades". International Society of Genetic Genealogy (ISOGG). Retrieved 8 January 2011.
- Joseph, Tony (2017), "How genetics is settling the Aryan migration debate", The Hindu
- Kivisild, T; Rootsi, S; Metspalu, M; Mastana, S; Kaldma, K; Parik, J; Metspalu, E; Adojaan, M; et al. (2003). "The Genetic Heritage of the Earliest Settlers Persists Both in Indian Tribal and Caste Populations". AJHG. 72 (2): 313–32. PMC 379225 . PMID 12536373. doi:10.1086/346068.
- Krahn, Thomas; FTDNA; Genetic Genealogy Community. "Family Tree DNA Draft Y-Chromosome Tree".
- Lazaridis, Iosif (2016), "The genetic structure of the world's first farmers", bioRxiv 059311
- Mascarenhas, Desmond D.; Raina, Anupuma; Aston, Christopher E.; Sanghera, Dharambir K. (2015), "Genetic and Cultural Reconstruction of the Migration of an Ancient Lineage", BioMed Research International, 2015, Article ID 651415, 16 pages: 1–16, doi:10.1155/2015/651415
- Mirabal, Sheyla; Regueiro, M; Cadenas, AM; Cavalli-Sforza, LL; Underhill, PA; Verbenko, DA; Limborska, SA; Herrera, RJ; et al. (2009). "Y-Chromosome distribution within the geo-linguistic landscape of northwestern Russia". European Journal of Human Genetics. 17 (10): 1260–1273. PMC 2986641 . PMID 19259129. doi:10.1038/ejhg.2009.6.
- Pamjav, Horolma; Fehér, Tibor; Németh, Endre; Pádár, Zsolt (2012), "Brief communication: new Y-chromosome binary markers improve phylogenetic resolution within haplogroup R1a1", American Journal of Physical Anthropology, 149 (4): 611–615, PMID 23115110, doi:10.1002/ajpa.22167
- Parpola, Asko (2015), The Roots of Hinduism. The early Aryans and the Indus Civilisation, Oxford University Press
- Poznik (2016), "Punctuated bursts in human male demography inferred from 1,244 worldwide Y-chromosome sequences", Nature Genetics, 48: 593–599, PMC 4884158 , PMID 27111036, doi:10.1038/ng.3559
- Regueiro, M; Cadenas, AM; Gayden, T; Underhill, PA; Herrera, RJ (2006). "Iran: Tricontinental Nexus for Y-Chromosome Driven Migration" (PDF). Hum Hered. 61 (3): 132–143. PMID 16770078. doi:10.1159/000093774.
- Sahoo, S; Singh, A; Himabindu, G; Banerjee, J; Sitalaximi, T; Gaikwad, S; Trivedi, R; Endicott, P; et al. (2006). "A prehistory of Indian Y chromosomes: Evaluating demic diffusion scenarios". Proceedings of the National Academy of Sciences. 103 (4): 843–848. Bibcode:2006PNAS..103..843S. PMC 1347984 . PMID 16415161. doi:10.1073/pnas.0507714103.
- Semenov, Alexander S.; Bulat, Vladimir V. (2016), "Ancient Paleo-DNA of Pre-Copper Age North-Eastern Europe: Establishing the Migration Traces of R1a1 Y-DNA Haplogroup", European Journal of Molecular Biotechnology, 11 (1): 40–54, doi:10.13187/ejmb.2016.11.40
- Semino, O; Passarino G, Oefner PJ (2000), "The genetic legacy of Paleolithic Homo sapiens sapiens in extant Europeans: A Y chromosome perspective", Science. 2000;290:1155–1159
- Sengupta, S; Zhivotovsky, LA; King, R; Mehdi, SQ; Edmonds, CA; Chow, CE; Lin, AA; Mitra, M; et al. (2006). "Polarity and Temporality of High-Resolution Y-Chromosome Distributions in India Identify Both Indigenous and Exogenous Expansions and Reveal Minor Genetic Influence of Central Asian Pastoralists". American Journal of Human Genetics. 78 (2): 202–21. PMC 1380230 . PMID 16400607. doi:10.1086/499411.
- Sharma, S; Rai, E; Sharma, P; Jena, M; Singh, S; Darvishi, K; Bhat, AK; Bhanwer, AJ; et al. (2009). "The Indian origin of paternal haplogroup R1a1(*)substantiates the autochthonous origin of Brahmins and the caste system". Journal of Human Genetics. 54 (1): 47–55. PMID 19158816. doi:10.1038/jhg.2008.2.
- Silva, Marina; et al. (2017), "A genetic chronology for the Indian Subcontinent points to heavily sex-biased dispersals", BMC Evolutionary Biology 2017 17:88, 17, doi:10.1186/s12862-017-0936-9
- Semino, O.; Passarino, G; Oefner, PJ; Lin, AA; Arbuzova, S; Beckman, LE; De Benedictis, G; Francalacci, P; et al. (2000). "The Genetic Legacy of Paleolithic Homo sapiens sapiens in Extant Europeans: A Y Chromosome Perspective" (PDF). Science. 290 (5494): 1155–59. Bibcode:2000Sci...290.1155S. PMID 11073453. doi:10.1126/science.290.5494.1155.. Copy can be found at http://www.historyofmacedonia.org/ConciseMacedonia/Y_Hromosomes.pdf.
- Thangaraj, Kumarasamy; Naidu, B. Prathap; Crivellaro, Federica; Tamang, Rakesh; Upadhyay, Shashank; Sharma, Varun Kumar; Reddy, Alla G.; Walimbe, S. R.; Chaubey, Gyaneshwer; Kivisild, T; Singh, L; et al. (2010). Cordaux, Richard, ed. "The Influence of Natural Barriers in Shaping the Genetic Structure of Maharashtra Populations". PLoS ONE. 5 (12): e15283. Bibcode:2010PLoSO...515283T. PMC 3004917 . PMID 21187967. doi:10.1371/journal.pone.0015283.
- Trautmann, Thomas (2005), The Aryan Debate, Oxford University Press
- Underhill, Peter A; Myres, Natalie M; Rootsi, Siiri; Metspalu, Mait; Zhivotovsky, Lev A; King, Roy J; Lin, Alice A; Chow, Cheryl-Emiliane T; et al. (2009). "Separating the post-Glacial coancestry of European and Asian Y chromosomes within haplogroup R1a". European Journal of Human Genetics. 18 (4): 479–84. PMC 2987245 . PMID 19888303. doi:10.1038/ejhg.2009.194.
- Underhill, Peter A. (2014), "The phylogenetic and geographic structure of Y-chromosome haplogroup R1a" (PDF), European Journal of Human Genetics, 23 (1): 124–131, ISSN 1018-4813, PMC 4266736 , PMID 24667786, doi:10.1038/ejhg.2014.50, retrieved 15 June 2016
- Underhill, Peter A. (2015), "The phylogenetic and geographic structure of Y-chromosome haplogroup R1a", European Journal of Human Genetics, 23: 124–131, PMC 4266736 , PMID 24667786, doi:10.1038/ejhg.2014.50
- Wells, R.S. (2001), "The Eurasian Heartland: A continental perspective on Y-chromosome diversity", Proceedings of the National Academy of Sciences of the USA, 98 (18): 10244–10249, PMC 56946 , PMID 11526236, doi:10.1073/pnas.171305098
- Y Chromosome Consortium "YCC" (2002). "A Nomenclature System for the Tree of Human Y-Chromosomal Binary Haplogroups". Genome Research. 12 (2): 339–348. PMC 155271 . PMID 11827954. doi:10.1101/gr.217602.
Further reading
| Extended content |
|---|
|
External links
| Wikimedia Commons has media related to Haplogroup R1a of Y-DNA. |
- DNA Tree
- FTDNA R1a Y-chromosome Haplogroup Project
- R1a1a1 and Subclades Y-DNA Project – Background Family Tree DNA R1a1a1
- TMRCA
- Various
- Danish Demes Regional DNA Project: Y-DNA Haplogroup R1a
- Eurogenes Blog, The Poltovka outlier
- Avotaynu Online, The Y-DNA Fingerprint of the Shpoler Zeida, a Tzaddik Who Touched the World