Glycogen


.tif.jpg)
Glycogen is a multibranched polysaccharide of glucose that serves as a form of energy storage in humans, animals,[2] fungi, and bacteria. The polysaccharide structure represents the main storage form of glucose in the body.
In humans, glycogen is made and stored primarily in the cells of the liver and the muscles, hydrated with three or four parts of water.[3] Glycogen functions as the secondary long-term energy storage, with the primary energy stores being fats held in adipose tissue. Muscle glycogen is converted into glucose by muscle cells, and liver glycogen converts to glucose for use throughout the body including the central nervous system.
Glycogen is the analogue of starch, a glucose polymer that functions as energy storage in plants. It has a structure similar to amylopectin (a component of starch), but is more extensively branched and compact than starch. Both are white powders in their dry state. Glycogen is found in the form of granules in the cytosol/cytoplasm in many cell types, and plays an important role in the glucose cycle. Glycogen forms an energy reserve that can be quickly mobilized to meet a sudden need for glucose, but one that is less compact than the energy reserves of triglycerides (lipids).
In the liver, glycogen can make up from 5–6% of the organ's fresh weight (100–120 g in an adult).[4] Only the glycogen stored in the liver can be made accessible to other organs. In the muscles, glycogen is found in a low concentration (1–2% of the muscle mass). The amount of glycogen stored in the body—especially within the muscles, liver, and red blood cells[5][6][7]—mostly depends on physical training, basal metabolic rate, and eating habits. Small amounts of glycogen are found in the kidneys, and even smaller amounts in certain glial cells in the brain and white blood cells. The uterus also stores glycogen during pregnancy to nourish the embryo.[8]
Structure


Glycogen is a branched biopolymer consisting of linear chains of glucose residues with further chains branching off every 8 to 12 glucoses or so. Glucoses are linked together linearly by α(1→4) glycosidic bonds from one glucose to the next. Branches are linked to the chains from which they are branching off by α(1→6) glycosidic bonds between the first glucose of the new branch and a glucose on the stem chain.[9]
Due to the way glycogen is synthesised, every glycogen granule has at its core a glycogenin protein.[10]
Glycogen in muscle, liver, and fat cells is stored in a hydrated form, composed of three or four parts of water per part of glycogen associated with 0.45 millimoles of potassium per gram of glycogen.[3]
Functions
Liver
As a meal containing carbohydrates or protein is eaten and digested, blood glucose levels rise, and the pancreas secretes insulin. Blood glucose from the portal vein enters liver cells (hepatocytes). Insulin acts on the hepatocytes to stimulate the action of several enzymes, including glycogen synthase. Glucose molecules are added to the chains of glycogen as long as both insulin and glucose remain plentiful. In this postprandial or "fed" state, the liver takes in more glucose from the blood than it releases.
After a meal has been digested and glucose levels begin to fall, insulin secretion is reduced, and glycogen synthesis stops. When it is needed for energy, glycogen is broken down and converted again to glucose. Glycogen phosphorylase is the primary enzyme of glycogen breakdown. For the next 8–12 hours, glucose derived from liver glycogen is the primary source of blood glucose used by the rest of the body for fuel.
Glucagon, another hormone produced by the pancreas, in many respects serves as a countersignal to insulin. In response to insulin levels being below normal (when blood levels of glucose begin to fall below the normal range), glucagon is secreted in increasing amounts and stimulates both glycogenolysis (the breakdown of glycogen) and gluconeogenesis (the production of glucose from other sources).
Muscle
Muscle cell glycogen appears to function as an immediate reserve source of available glucose for muscle cells. Other cells that contain small amounts use it locally, as well. As muscle cells lack glucose-6-phosphatase, which is required to pass glucose into the blood, the glycogen they store is available solely for internal use and is not shared with other cells. This is in contrast to liver cells, which, on demand, readily do break down their stored glycogen into glucose and send it through the blood stream as fuel for other organs.
History
Glycogen was discovered by Claude Bernard. His experiments showed that the liver contained a substance that could give rise to reducing sugar by the action of a "ferment" in the liver. By 1857, he described the isolation of a substance he called "la matière glycogène", or "sugar-forming substance". Soon after the discovery of glycogen in the liver, A. Sanson found that muscular tissue also contains glycogen. The empirical formula for glycogen of (C
6H
10O
5)n was established by Kekule in 1858.[11]
Metabolism
Synthesis
Glycogen synthesis is, unlike its breakdown, endergonic—it requires the input of energy. Energy for glycogen synthesis comes from uridine triphosphate (UTP), which reacts with glucose-1-phosphate, forming UDP-glucose, in a reaction catalysed by UTP—glucose-1-phosphate uridylyltransferase. Glycogen is synthesized from monomers of UDP-glucose initially by the protein glycogenin, which has two tyrosine anchors for the reducing end of glycogen, since glycogenin is a homodimer. After about eight glucose molecules have been added to a tyrosine residue, the enzyme glycogen synthase progressively lengthens the glycogen chain using UDP-glucose, adding α(1→4)-bonded glucose. The glycogen branching enzyme catalyzes the transfer of a terminal fragment of six or seven glucose residues from a nonreducing end to the C-6 hydroxyl group of a glucose residue deeper into the interior of the glycogen molecule. The branching enzyme can act upon only a branch having at least 11 residues, and the enzyme may transfer to the same glucose chain or adjacent glucose chains.
Breakdown
Glycogen is cleaved from the nonreducing ends of the chain by the enzyme glycogen phosphorylase to produce monomers of glucose-1-phosphate:
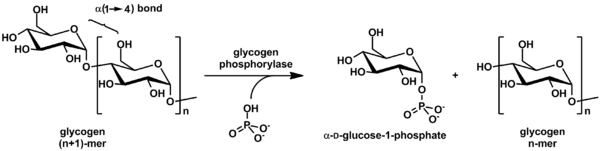
In vivo, phosphorolysis proceeds in the direction of glycogen breakdown because the ratio of phosphate and glucose-1-phosphate is usually greater than 100.[12] Glucose-1-phosphate is then converted to glucose 6-phosphate (G6P) by phosphoglucomutase. A special debranching enzyme is needed to remove the α(1-6) branches in branched glycogen and reshape the chain into a linear polymer. The G6P monomers produced have three possible fates:
- G6P can continue on the glycolysis pathway and be used as fuel.
- G6P can enter the pentose phosphate pathway via the enzyme glucose-6-phosphate dehydrogenase to produce NADPH and 5-carbon sugars.
- In the liver and kidney, G6P can be dephosphorylated back to glucose by the enzyme glucose 6-phosphatase. This is the final step in the gluconeogenesis pathway.
Clinical relevance
Disorders of glycogen metabolism
The most common disease in which glycogen metabolism becomes abnormal is diabetes, in which, because of abnormal amounts of insulin, liver glycogen can be abnormally accumulated or depleted. Restoration of normal glucose metabolism usually normalizes glycogen metabolism, as well.
In hypoglycemia caused by excessive insulin, liver glycogen levels are high, but the high insulin levels prevent the glycogenolysis necessary to maintain normal blood sugar levels. Glucagon is a common treatment for this type of hypoglycemia.
Various inborn errors of metabolism are caused by deficiencies of enzymes necessary for glycogen synthesis or breakdown. These are collectively referred to as glycogen storage diseases.
Glycogen depletion and endurance exercise
Long-distance athletes, such as marathon runners, cross-country skiers, and cyclists, often experience glycogen depletion, where almost all of the athlete's glycogen stores are depleted after long periods of exertion without sufficient carbohydrate consumption. This phenomenon is referred to as "hitting the wall".
Glycogen depletion can be forestalled in three possible ways. First, during exercise, carbohydrates with the highest possible rate of conversion to blood glucose (high glycemic index) are ingested continuously. The best possible outcome of this strategy replaces about 35% of glucose consumed at heart rates above about 80% of maximum. Second, through endurance training adaptations and specialized regimens (e.g. fasted low-intensity endurance training), the body can condition type I muscle fibers to improve both fuel use efficiency and workload capacity to increase the percentage of fatty acids used as fuel,[13][14] sparing carbohydrate use from all sources. Third, by consuming large quantities of carbohydrates after depleting glycogen stores as a result of exercise or diet, the body can increase storage capacity of intramuscular glycogen stores.[15][16][17] This process is known as carbohydrate loading. In general, glycemic index of carbohydrate source does not matter since muscular insulin sensitivity is increased as a result of temporary glycogen depletion.[18][19]
When experiencing glycogen debt, athletes often experience extreme fatigue to the point that it is difficult to move. As a reference, the very best professional cyclists in the world will usually finish a 4- to 5-hr stage race right at the limit of glycogen depletion using the first three strategies.
When athletes ingest both carbohydrate and caffeine following exhaustive exercise, their glycogen stores tend to be replenished more rapidly;[20][21][22] however, the minimum dose of caffeine at which there is a clinically significant effect on glycogen repletion has not been established.[22]
See also
References
- ↑ William D. McArdle; Frank I. Katch; Victor L. Katch (2006). Exercise physiology: energy, nutrition, and human performance (6 ed.). Lippincott Williams & Wilkins. p. 12. ISBN 978-0-7817-4990-9.
- ↑ Sadava; et al. (2011). Life (9th, International ed.). W. H. Freeman. ISBN 9781429254311.
- 1 2 Kreitzman SN, Coxon AY, Szaz KF (1992). "Glycogen storage: illusions of easy weight loss, excessive weight regain, and distortions in estimates of body composition" (PDF). The American Journal of Clinical Nutrition. 56 (1 Suppl): 292s–293s. PMID 1615908.
- ↑ Guyton, Arthur C.; John Edward Hall (2011). Guyton and Hall Textbook of Medical Physiology. New York, New York: Saunders/Elsevier. ISBN 978-5-98657-013-6.
- ↑ Moses SW, Bashan N, Gutman A (December 1972). "Glycogen metabolism in the normal red blood cell". Blood. 40 (6): 836–43. PMID 5083874.
- ↑ Ingermann RL, Virgin GL (1987). "Glycogen content and release of glucose from red blood cells of the sipunculan worm themiste dyscrita" (PDF). J Exp Biol. 129: 141–9.
- ↑ Miwa I, Suzuki S (November 2002). "An improved quantitative assay of glycogen in erythrocytes". Annals of Clinical Biochemistry. 39 (Pt 6): 612–3. PMID 12564847. doi:10.1258/000456302760413432.
- ↑ Campbell, Neil A.; Brad Williamson; Robin J. Heyden (2006). Biology: Exploring Life. Boston: Pearson Prentice Hall. ISBN 0-13-250882-6.
- ↑ Berg, Tymoczko & Stryer (2012). Biochemistry (7th, International ed.). W. H. Freeman. p. 338. ISBN 1429203145.
- ↑ Berg; et al. (2012). Biochemistry (7th, International ed.). W. H. Freeman. p. 650.
- ↑ F. G. Young (1957). "Claude Bernard and the Discovery of Glycogen". British Medical Journal. 1 (5033 (Jun. 22, 1957)): 1431–7. JSTOR 25382898. doi:10.1136/bmj.1.5033.1431.
- ↑ Stryer, L. (1988) Biochemistry, 3rd ed., Freeman (p. 451)
- ↑ http://www.bodyrecomposition.com/training/methods-of-endurance-training-part-1.html
- ↑ http://www.bodyrecomposition.com/fat-loss/qa-steady-state-vs-tempo-training-and-fat-loss.html
- ↑ McDonald, Lyle. "Research Review: An In-Depth Look Into Carbing Up On The Cyclical Ketogenic Diet". Retrieved 19 February 2017.
- ↑ McDonald, Lyle. The Ketogenic Diet: A Complete Guide for the Dieter and the Practitioner. Lyle McDonald, 1998
- ↑ "Costill DL et. al. Muscle glycogen utilization during prolonged exercise on successive days. J Appl Physiol (1971) 31: 834-838."
- ↑ Glycogen depletion and increased insulin sensitivity and responsiveness in muscle after exerciseAm J Physiol Endocrinol MetabDecember 1, 1986 251:(6) E664-E669
- ↑ McDonald, Lyle. The Ultimate Diet 2.0. Lyle McDonald, 2003
- ↑ Pedersen DJ, Lessard SJ, Coffey VG, et al. (July 2008). "High rates of muscle glycogen resynthesis after exhaustive exercise when carbohydrate is coingested with caffeine". Journal of Applied Physiology (Original article). 105 (1): 7–13. PMID 18467543. doi:10.1152/japplphysiol.01121.2007.
- ↑ Gaudet, Laura; Jackson, Allen; Streitz, Carmyn; McIntire, Kyle; McDaniel, Larry. "The Effects Of Caffeine On Athletic Performance". Clute Institute. Retrieved 17 June 2014.
- 1 2 Beelen M, Burke LM, Gibala MJ, van Loon L JC (December 2010). "Nutritional strategies to promote postexercise recovery". International Journal of Sport Nutrition and Exercise Metabolism. 20 (6): 515–532. PMID 21116024. doi:10.1123/ijsnem.20.6.515.
External links
- Glycogen detection using Periodic Acid Schiff Staining
- Glycogen storage disease – McArdle's Disease Website
- Glycogen at the US National Library of Medicine Medical Subject Headings (MeSH)