DNA-binding protein


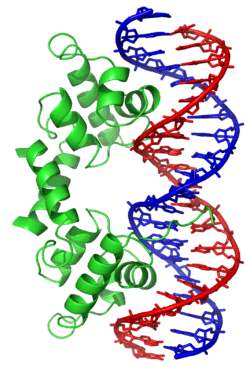

DNA-binding proteins are proteins composed of DNA-binding domains and thus have a specific or general affinity for either single or double stranded DNA.[3][4][5] Sequence-specific DNA-binding proteins generally interact with the major groove of B-DNA, because it exposes more functional groups that identify a base pair. However, there are some known minor groove DNA-binding ligands such as netropsin,[6] distamycin, Hoechst 33258, pentamidine, DAPI and others.[7]
Examples
DNA-binding proteins include transcription factors which modulate the process of transcription, various polymerases, nucleases which cleave DNA molecules, and histones which are involved in chromosome packaging and transcription in the cell nucleus. DNA-binding proteins can incorporate such domains as the zinc finger, the helix-turn-helix, and the leucine zipper (among many others) that facilitate binding to nucleic acid. There are also more unusual examples such as transcription activator like effectors.
Non-specific DNA-protein interactions
Structural proteins that bind DNA are well-understood examples of non-specific DNA-protein interactions. Within chromosomes, DNA is held in complexes with structural proteins. These proteins organize the DNA into a compact structure called chromatin. In eukaryotes, this structure involves DNA binding to a complex of small basic proteins called histones. In prokaryotes, multiple types of proteins are involved.[8][9] The histones form a disk-shaped complex called a nucleosome, which contains two complete turns of double-stranded DNA wrapped around its surface. These non-specific interactions are formed through basic residues in the histones making ionic bonds to the acidic sugar-phosphate backbone of the DNA, and are therefore largely independent of the base sequence.[10] Chemical modifications of these basic amino acid residues include methylation, phosphorylation and acetylation.[11] These chemical changes alter the strength of the interaction between the DNA and the histones, making the DNA more or less accessible to transcription factors and changing the rate of transcription.[12] Other non-specific DNA-binding proteins in chromatin include the high-mobility group (HMG) proteins, which bind to bent or distorted DNA.[13] Biophysical studies show that these architectural HMG proteins bind, bend and loop DNA to perform its biological functions.[14][15] These proteins are important in bending arrays of nucleosomes and arranging them into the larger structures that form chromosomes.[16]
DNA-binding proteins that specifically bind single-stranded DNA
A distinct group of DNA-binding proteins are the DNA-binding proteins that specifically bind single-stranded DNA. In humans, replication protein A is the best-understood member of this family and is used in processes where the double helix is separated, including DNA replication, recombination and DNA repair.[17] These binding proteins seem to stabilize single-stranded DNA and protect it from forming stem-loops or being degraded by nucleases.
Binding to specific DNA sequences
In contrast, other proteins have evolved to bind to specific DNA sequences. The most intensively studied of these are the various transcription factors, which are proteins that regulate transcription. Each transcription factor binds to one specific set of DNA sequences and activates or inhibits the transcription of genes that have these sequences near their promoters. The transcription factors do this in two ways. Firstly, they can bind the RNA polymerase responsible for transcription, either directly or through other mediator proteins; this locates the polymerase at the promoter and allows it to begin transcription.[18] Alternatively, transcription factors can bind enzymes that modify the histones at the promoter. This alters the accessibility of the DNA template to the polymerase.[19]
These DNA targets can occur throughout an organism's genome. Thus, changes in the activity of one type of transcription factor can affect thousands of genes.[20] Thus, these proteins are often the targets of the signal transduction processes that control responses to environmental changes or cellular differentiation and development. The specificity of these transcription factors' interactions with DNA come from the proteins making multiple contacts to the edges of the DNA bases, allowing them to read the DNA sequence. Most of these base-interactions are made in the major groove, where the bases are most accessible.[21] Mathematical descriptions of protein-DNA binding taking into account sequence-specificity, and competitive and cooperative binding of proteins of different types are usually performed with the help of the lattice models.[22] Computational methods to identify the DNA binding sequence specificity have been proposed to make a good use of the abundant sequence data in the post-genomic era.[23]
See also
- bZIP domain
- Comparison of nucleic acid simulation software
- DNA-binding domain
- Helix-loop-helix
- Helix-turn-helix
- HMG-box
- Leucine zipper
- Lexitropsin (a semi-synthetic DNA-binding ligand)
- Single-strand binding protein
- Zinc finger
References
- ↑ Created from PDB 1LMB
- ↑ Created from PDB 1RVA
- ↑ Travers, A. A. (1993). DNA-protein interactions. London: Springer. ISBN 978-0-412-25990-6.
- ↑ Pabo CO, Sauer RT (1984). "Protein-DNA recognition". Annu. Rev. Biochem. 53 (1): 293–321. PMID 6236744. doi:10.1146/annurev.bi.53.070184.001453.
- ↑ Dickerson R.E. (1983). "The DNA helix and how it is read". Sci Am. 249 (6): 94–111. doi:10.1038/scientificamerican1283-94.
- ↑ Zimmer C, Wähnert U (1986). "Nonintercalating DNA-binding ligands: specificity of the interaction and their use as tools in biophysical, biochemical and biological investigations of the genetic material". Prog. Biophys. Mol. Biol. 47 (1): 31–112. PMID 2422697. doi:10.1016/0079-6107(86)90005-2.
- ↑ Dervan PB (April 1986). "Design of sequence-specific DNA-binding molecules". Science. 232 (4749): 464–71. PMID 2421408. doi:10.1126/science.2421408.
- ↑ Sandman K, Pereira S, Reeve J (1998). "Diversity of prokaryotic chromosomal proteins and the origin of the nucleosome". Cell Mol Life Sci. 54 (12): 1350–64. PMID 9893710. doi:10.1007/s000180050259.
- ↑ Dame RT (2005). "The role of nucleoid-associated proteins in the organization and compaction of bacterial chromatin". Mol. Microbiol. 56 (4): 858–70. PMID 15853876. doi:10.1111/j.1365-2958.2005.04598.x.
- ↑ Luger K, Mäder A, Richmond R, Sargent D, Richmond T (1997). "Crystal structure of the nucleosome core particle at 2.8 A resolution". Nature. 389 (6648): 251–60. PMID 9305837. doi:10.1038/38444.
- ↑ Jenuwein T, Allis C (2001). "Translating the histone code". Science. 293 (5532): 1074–80. PMID 11498575. doi:10.1126/science.1063127.
- ↑ Ito T (2003). "Nucleosome assembly and remodelling". Curr Top Microbiol Immunol. 274: 1–22. PMID 12596902. doi:10.1007/978-3-642-55747-7_1.
- ↑ Thomas J (2001). "HMG1 and 2: architectural DNA-binding proteins". Biochem Soc Trans. 29 (Pt 4): 395–401. PMID 11497996. doi:10.1042/BST0290395.
- ↑ D. Murugesapillai et al, DNA bridging and looping by HMO1 provides a mechanism for stabilizing nucleosome-free chromatin, Nucl Acids Res (2014) 42 (14): 8996-9004
- ↑ D. Murugesapillai et al, Single-molecule studies of high-mobility group B architectural DNA bending proteins, Biophys Rev (2016) doi:10.1007/s12551-016-0236-4
- ↑ Grosschedl R, Giese K, Pagel J (1994). "HMG domain proteins: architectural elements in the assembly of nucleoprotein structures". Trends Genet. 10 (3): 94–100. PMID 8178371. doi:10.1016/0168-9525(94)90232-1.
- ↑ Iftode C, Daniely Y, Borowiec J (1999). "Replication protein A (RPA): the eukaryotic SSB". Crit Rev Biochem Mol Biol. 34 (3): 141–80. PMID 10473346. doi:10.1080/10409239991209255.
- ↑ Myers L, Kornberg R (2000). "Mediator of transcriptional regulation". Annu Rev Biochem. 69 (1): 729–49. PMID 10966474. doi:10.1146/annurev.biochem.69.1.729.
- ↑ Spiegelman B, Heinrich R (2004). "Biological control throughs regulated transcriptional coactivators". Cell. 119 (2): 157–67. PMID 15479634. doi:10.1016/j.cell.2004.09.037.
- ↑ Li Z, Van Calcar S, Qu C, Cavenee W, Zhang M, Ren B (2003). "A global transcriptional regulatory role for c-Myc in Burkitt's lymphoma cells". Proc Natl Acad Sci USA. 100 (14): 8164–9. PMC 166200
 . PMID 12808131. doi:10.1073/pnas.1332764100.
. PMID 12808131. doi:10.1073/pnas.1332764100. - ↑ Pabo C, Sauer R (1984). "Protein-DNA recognition". Annu Rev Biochem. 53 (1): 293–321. PMID 6236744. doi:10.1146/annurev.bi.53.070184.001453.
- ↑ Teif V.B.; Rippe K. (2010). "Statistical-mechanical lattice models for protein-DNA binding in chromatin.". Journal of Physics: Condensed Matter. 22 (41): 414105. PMID 21386588. arXiv:1004.5514 . doi:10.1088/0953-8984/22/41/414105.
- ↑ Wong KC, Chan TM, Peng C., Li Y., and Zhang Z. "DNA Motif Elucidation using belief propagation" Nucleic Acids Research Advanced Online June 2013; doi:10.1093/nar/gkt574 PMID 23814189
External links
- Abalone tool for modeling DNA-ligand interactions.
- DBD database of predicted transcription factors Uses a curated set of DNA-binding domains to predict transcription factors in all completely sequenced genomes
- DNA-Binding Proteins at the US National Library of Medicine Medical Subject Headings (MeSH)