Amino acid

Amino acids are organic compounds containing amine (-NH2) and carboxyl (-COOH) functional groups, along with a side chain (R group) specific to each amino acid.[1][2][3] The key elements of an amino acid are carbon, hydrogen, oxygen, and nitrogen, although other elements are found in the side chains of certain amino acids. About 500 amino acids are known (though only 20 appear in the genetic code) and can be classified in many ways.[4] They can be classified according to the core structural functional groups' locations as alpha- (α-), beta- (β-), gamma- (γ-) or delta- (δ-) amino acids; other categories relate to polarity, pH level, and side chain group type (aliphatic, acyclic, aromatic, containing hydroxyl or sulfur, etc.). In the form of proteins, amino acid residues form the second-largest component (water is the largest) of human muscles and other tissues.[5] Beyond their role as residues in proteins, amino acids participate in a number of processes such as neurotransmitter transport and biosynthesis.
In biochemistry, amino acids having both the amine and the carboxylic acid groups attached to the first (alpha-) carbon atom have particular importance. They are known as 2-, alpha-, or α-amino acids (generic formula H2NCHRCOOH in most cases,[6] where R is an organic substituent known as a "side chain");[7] often the term "amino acid" is used to refer specifically to these. They include the 22 proteinogenic ("protein-building") amino acids,[8][9][10] which combine into peptide chains ("polypeptides") to form the building-blocks of a vast array of proteins.[11] These are all L-stereoisomers ("left-handed" isomers), although a few D-amino acids ("right-handed") occur in bacterial envelopes, as a neuromodulator (D-serine), and in some antibiotics.[12]
Twenty of the proteinogenic amino acids are encoded directly by triplet codons in the genetic code and are known as "standard" amino acids. The other two ("non-standard" or "non-canonical") are selenocysteine (present in many prokaryotes as well as most eukaryotes, but not coded directly by DNA), and pyrrolysine (found only in some archea and one bacterium). Pyrrolysine and selenocysteine are encoded via variant codons; for example, selenocysteine is encoded by stop codon and SECIS element.[13][14][15] N-formylmethionine (which is often the initial amino acid of proteins in bacteria, mitochondria, and chloroplasts) is generally considered as a form of methionine rather than as a separate proteinogenic amino acid. Codon–tRNA combinations not found in nature can also be used to "expand" the genetic code and form novel proteins known as alloproteins incorporating non-proteinogenic amino acids.[16][17][18]
Many important proteinogenic and non-proteinogenic amino acids have biological functions. For example, in the human brain, glutamate (standard glutamic acid) and gamma-amino-butyric acid ("GABA", non-standard gamma-amino acid) are, respectively, the main excitatory and inhibitory neurotransmitters.[19] Hydroxyproline, a major component of the connective tissue collagen, is synthesised from proline. Glycine is a biosynthetic precursor to porphyrins used in red blood cells. Carnitine is used in lipid transport.
Nine proteinogenic amino acids are called "essential" for humans because they cannot be produced from other compounds by the human body and so must be taken in as food. Others may be conditionally essential for certain ages or medical conditions. Essential amino acids may also differ between species.[20]
Because of their biological significance, amino acids are important in nutrition and are commonly used in nutritional supplements, fertilizers, and food technology. Industrial uses include the production of drugs, biodegradable plastics, and chiral catalysts.
History
The first few amino acids were discovered in the early 19th century. In 1806, French chemists Louis-Nicolas Vauquelin and Pierre Jean Robiquet isolated a compound in asparagus that was subsequently named asparagine, the first amino acid to be discovered.[21][22] Cystine was discovered in 1810,[23] although its monomer, cysteine, remained undiscovered until 1884.[22][24] Glycine and leucine were discovered in 1820.[25] The last of the 20 common amino acids to be discovered was threonine in 1935 by William Cumming Rose, who also determined the essential amino acids and established the minimum daily requirements of all amino acids for optimal growth.[26][27]
Usage of the term amino acid in the English language is from 1898.[28] Proteins were found to yield amino acids after enzymatic digestion or acid hydrolysis. In 1902, Emil Fischer and Franz Hofmeister proposed that proteins are the result of the formation of bonds between the amino group of one amino acid with the carboxyl group of another, in a linear structure that Fischer termed "peptide".[29]
General structure
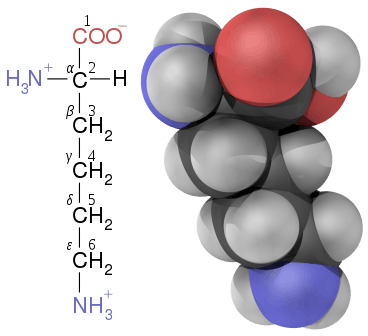

In the structure shown at the top of the page, R represents a side chain specific to each amino acid. The carbon atom next to the carboxyl group (which is therefore numbered 2 in the carbon chain starting from that functional group) is called the α–carbon. Amino acids containing an amino group bonded directly to the alpha carbon are referred to as alpha amino acids.[30] These include amino acids such as proline which contain secondary amines, which used to be often referred to as "imino acids".[31][32][33]
Isomerism
The alpha amino acids are the most common form found in nature, but only when occurring in the L-isomer. The alpha carbon is a chiral carbon atom, with the exception of glycine which has two indistinguishable hydrogen atoms on the alpha carbon.[34] Therefore, all alpha amino acids but glycine can exist in either of two enantiomers, called L or D amino acids, which are mirror images of each other (see also Chirality). While L-amino acids represent all of the amino acids found in proteins during translation in the ribosome, D-amino acids are found in some proteins produced by enzyme posttranslational modifications after translation and translocation to the endoplasmic reticulum, as in exotic sea-dwelling organisms such as cone snails.[35] They are also abundant components of the peptidoglycan cell walls of bacteria,[36] and D-serine may act as a neurotransmitter in the brain.[37] D-amino acids are used in racemic crystallography to create centrosymmetric crystals, which (depending on the protein) may allow for easier and more robust protein structure determination.[38] The L and D convention for amino acid configuration refers not to the optical activity of the amino acid itself but rather to the optical activity of the isomer of glyceraldehyde from which that amino acid can, in theory, be synthesized (D-glyceraldehyde is dextrorotatory; L-glyceraldehyde is levorotatory). In alternative fashion, the (S) and (R) designators are used to indicate the absolute stereochemistry. Almost all of the amino acids in proteins are (S) at the α carbon, with cysteine being (R) and glycine non-chiral.[39] Cysteine has its side chain in the same geometric position as the other amino acids, but the R/S terminology is reversed because of the higher atomic number of sulfur compared to the carboxyl oxygen gives the side chain a higher priority, whereas the atoms in most other side chains give them lower priority.
Side chains
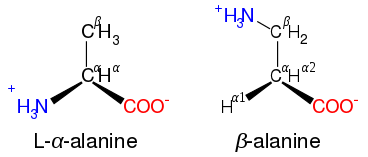
In amino acids that have a carbon chain attached to the α–carbon (such as lysine, shown to the right) the carbons are labeled in order as α, β, γ, δ, and so on.[40] In some amino acids, the amine group is attached to the β or γ-carbon, and these are therefore referred to as beta or gamma amino acids.
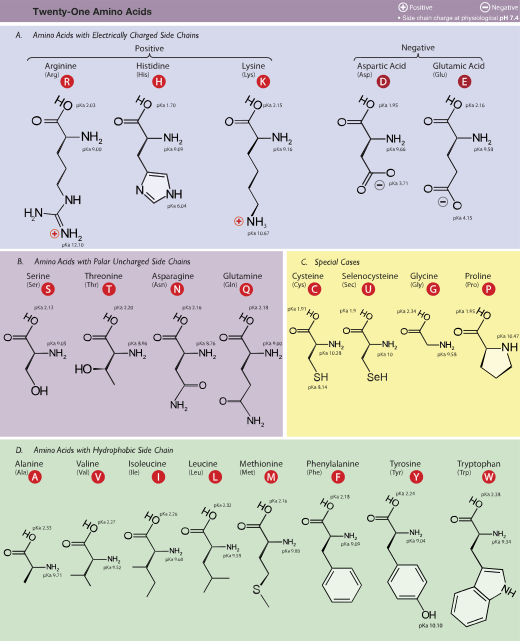
Amino acids are usually classified by the properties of their side chain into four groups. The side chain can make an amino acid a weak acid or a weak base, and a hydrophile if the side chain is polar or a hydrophobe if it is nonpolar.[34] The chemical structures of the 22 standard amino acids, along with their chemical properties, are described more fully in the article on these proteinogenic amino acids.
The phrase "branched-chain amino acids" or BCAA refers to the amino acids having aliphatic side chains that are non-linear; these are leucine, isoleucine, and valine. Proline is the only proteinogenic amino acid whose side-group links to the α-amino group and, thus, is also the only proteinogenic amino acid containing a secondary amine at this position.[34] In chemical terms, proline is, therefore, an imino acid, since it lacks a primary amino group,[41] although it is still classed as an amino acid in the current biochemical nomenclature,[42] and may also be called an "N-alkylated alpha-amino acid".[43]
Zwitterions
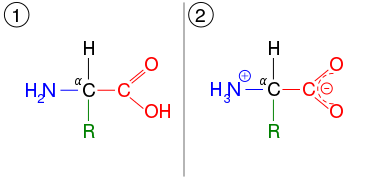
The α-carboxylic acid group of amino acids is a weak acid, meaning that it releases a hydron (such as a proton) at moderate pH values. In other words, carboxylic acid groups (−CO2H) can be deprotonated to become negative carboxylates (−CO2− ). The negatively charged carboxylate ion predominates at pH values greater than the pKa of the carboxylic acid group (mean for the 20 common amino acids is about 2.2, see the table of amino acid structures above). In a complementary fashion, the α-amine of amino acids is a weak base, meaning that it accepts a proton at moderate pH values. In other words, α-amino groups (NH2−) can be protonated to become positive α-ammonium groups (+NH3−). The positively charged α-ammonium group predominates at pH values less than the pKa of the α-ammonium group (mean for the 20 common α-amino acids is about 9.4).
Because all amino acids contain amine and carboxylic acid functional groups, they share amphiprotic properties.[34] Below pH 2.2, the predominant form will have a neutral carboxylic acid group and a positive α-ammonium ion (net charge +1), and above pH 9.4, a negative carboxylate and neutral α-amino group (net charge −1). But at pH between 2.2 and 9.4, an amino acid usually contains both a negative carboxylate and a positive α-ammonium group, as shown in structure (2) on the right, so has net zero charge. This molecular state is known as a zwitterion, from the German Zwitter meaning hermaphrodite or hybrid.[44] The fully neutral form (structure (1) on the left) is a very minor species in aqueous solution throughout the pH range (less than 1 part in 107). Amino acids exist as zwitterions also in the solid phase, and crystallize with salt-like properties unlike typical organic acids or amines.
Isoelectric point
The variation in titration curves when the amino acids are grouped by category can be seen here. With the exception of tyrosine, using titration to distinguish between hydrophobic amino acids is problematic.
At pH values between the two pKa values, the zwitterion predominates, but coexists in dynamic equilibrium with small amounts of net negative and net positive ions. At the exact midpoint between the two pKa values, the trace amount of net negative and trace of net positive ions exactly balance, so that average net charge of all forms present is zero.[45] This pH is known as the isoelectric point pI, so pI = ½(pKa1 + pKa2). The individual amino acids all have slightly different pKa values, so have different isoelectric points. For amino acids with charged side chains, the pKa of the side chain is involved. Thus for Asp, Glu with negative side chains, pI = ½(pKa1 + pKaR), where pKaR is the side chain pKa. Cysteine also has potentially negative side chain with pKaR = 8.14, so pI should be calculated as for Asp and Glu, even though the side chain is not significantly charged at neutral pH. For His, Lys, and Arg with positive side chains, pI = ½(pKaR + pKa2). Amino acids have zero mobility in electrophoresis at their isoelectric point, although this behaviour is more usually exploited for peptides and proteins than single amino acids. Zwitterions have minimum solubility at their isoelectric point and some amino acids (in particular, with non-polar side chains) can be isolated by precipitation from water by adjusting the pH to the required isoelectric point.
Occurrence and functions in biochemistry
Proteinogenic amino acids
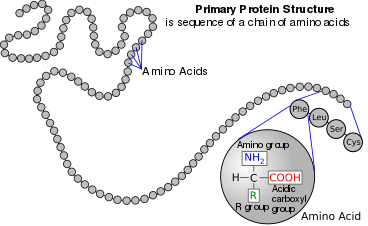
Amino acids are the structural units (monomers) that make up proteins. They join together to form short polymer chains called peptides or longer chains called either polypeptides or proteins. These polymers are linear and unbranched, with each amino acid within the chain attached to two neighboring amino acids. The process of making proteins is called translation and involves the step-by-step addition of amino acids to a growing protein chain by a ribozyme that is called a ribosome.[46] The order in which the amino acids are added is read through the genetic code from an mRNA template, which is a RNA copy of one of the organism's genes.
Twenty-two amino acids are naturally incorporated into polypeptides and are called proteinogenic or natural amino acids.[34] Of these, 20 are encoded by the universal genetic code. The remaining 2, selenocysteine and pyrrolysine, are incorporated into proteins by unique synthetic mechanisms. Selenocysteine is incorporated when the mRNA being translated includes a SECIS element, which causes the UGA codon to encode selenocysteine instead of a stop codon.[47] Pyrrolysine is used by some methanogenic archaea in enzymes that they use to produce methane. It is coded for with the codon UAG, which is normally a stop codon in other organisms.[48] This UAG codon is followed by a PYLIS downstream sequence.[49]
Non-proteinogenic amino acids
Aside from the 22 proteinogenic amino acids, many non-proteinogenic amino acids are known. Those either are not found in proteins (for example carnitine, GABA, Levothyroxine) or are not produced directly and in isolation by standard cellular machinery (for example, hydroxyproline and selenomethionine).
Non-proteinogenic amino acids that are found in proteins are formed by post-translational modification, which is modification after translation during protein synthesis. These modifications are often essential for the function or regulation of a protein. For example, the carboxylation of glutamate allows for better binding of calcium cations,[50] and collagen contains hydroxyproline, generated by hydroxylation of proline.[51] Another example is the formation of hypusine in the translation initiation factor EIF5A, through modification of a lysine residue.[52] Such modifications can also determine the localization of the protein, e.g., the addition of long hydrophobic groups can cause a protein to bind to a phospholipid membrane.[53]
Some non-proteinogenic amino acids are not found in proteins. Examples include 2-aminoisobutyric acid and the neurotransmitter gamma-aminobutyric acid. Non-proteinogenic amino acids often occur as intermediates in the metabolic pathways for standard amino acids – for example, ornithine and citrulline occur in the urea cycle, part of amino acid catabolism (see below).[54] A rare exception to the dominance of α-amino acids in biology is the β-amino acid beta alanine (3-aminopropanoic acid), which is used in plants and microorganisms in the synthesis of pantothenic acid (vitamin B5), a component of coenzyme A.[55]
D-amino acid natural abundance
D-isomers are uncommon in live organisms. For instance, gramicidin is a polypeptide made up from mixture of D- and L-amino acids.[56] Other compounds containing D-amino acid are tyrocidine and valinomycin. These compounds disrupt bacterial cell walls, particularly in Gram-positive bacteria. Only 837 D-amino acids were found in Swiss-Prot database (187 million amino acids analysed).[57]
Non-standard amino acids
The 20 amino acids that are encoded directly by the codons of the universal genetic code are called standard or canonical amino acids. A modified form of methionine (N-formylmethionine) is often incorporated in place of methionine as the initial amino acid of proteins in bacteria, mitochondria and chloroplasts. Other amino acids are called non-standard or non-canonical. Most of the non-standard amino acids are also non-proteinogenic (i.e. they cannot be incorporated into proteins during translation), but two of them are proteinogenic, as they can be incorporated translationally into proteins by exploiting information not encoded in the universal genetic code.
The two non-standard proteinogenic amino acids are selenocysteine (present in many non-eukaryotes as well as most eukaryotes, but not coded directly by DNA) and pyrrolysine (found only in some archaea and one bacterium). The incorporation of these non-standard amino acids is rare. For example, 25 human proteins include selenocysteine (Sec) in their primary structure,[58] and the structurally characterized enzymes (selenoenzymes) employ Sec as the catalytic moiety in their active sites.[59] Pyrrolysine and selenocysteine are encoded via variant codons. For example, selenocysteine is encoded by stop codon and SECIS element.[13][14][15]
In human nutrition
When taken up into the human body from the diet, the 20 standard amino acids either are used to synthesize proteins and other biomolecules or are oxidized to urea and carbon dioxide as a source of energy.[60] The oxidation pathway starts with the removal of the amino group by a transaminase; the amino group is then fed into the urea cycle. The other product of transamidation is a keto acid that enters the citric acid cycle.[61] Glucogenic amino acids can also be converted into glucose, through gluconeogenesis.[62] Of the 20 standard amino acids, nine (His, Ile, Leu, Lys, Met, Phe, Thr, Trp and Val) are called essential amino acids because the human body cannot synthesize them from other compounds at the level needed for normal growth, so they must be obtained from food.[63][64][65] In addition, cysteine, taurine, tyrosine, and arginine are considered semiessential amino-acids in children (though taurine is not technically an amino acid), because the metabolic pathways that synthesize these amino acids are not fully developed.[66][67] The amounts required also depend on the age and health of the individual, so it is hard to make general statements about the dietary requirement for some amino acids. Dietary exposure to the non-standard amino acid BMAA has been linked to human neurodegenerative diseases, including ALS.[68][69]
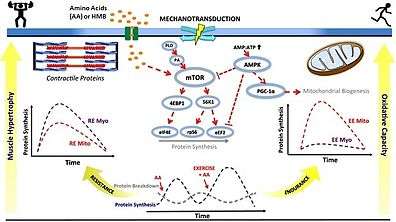
• PA: phosphatidic acid
• mTOR: mechanistic target of rapamycin
• AMP: adenosine monophosphate
• ATP: adenosine triphosphate
• AMPK: AMP-activated protein kinase
• PGC‐1α: peroxisome proliferator-activated receptor gamma coactivator-1α
• S6K1: p70S6 kinase
• 4EBP1: eukaryotic translation initiation factor 4E-binding protein 1
• eIF4E: eukaryotic translation initiation factor 4E
• RPS6: ribosomal protein S6
• eEF2: eukaryotic elongation factor 2
• RE: resistance exercise; EE: endurance exercise
• Myo: myofibrillar; Mito: mitochondrial
• AA: amino acids
• HMB: β-hydroxy β-methylbutyric acid
• ↑ represents activation
• Τ represents inhibition
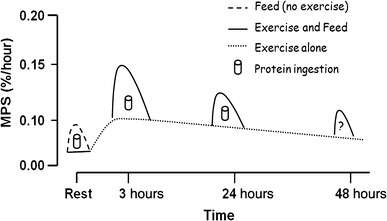
Non-protein functions
In humans, non-protein amino acids also have important roles as metabolic intermediates, such as in the biosynthesis of the neurotransmitter gamma-amino-butyric acid (GABA). Many amino acids are used to synthesize other molecules, for example:
- Tryptophan is a precursor of the neurotransmitter serotonin.[76]
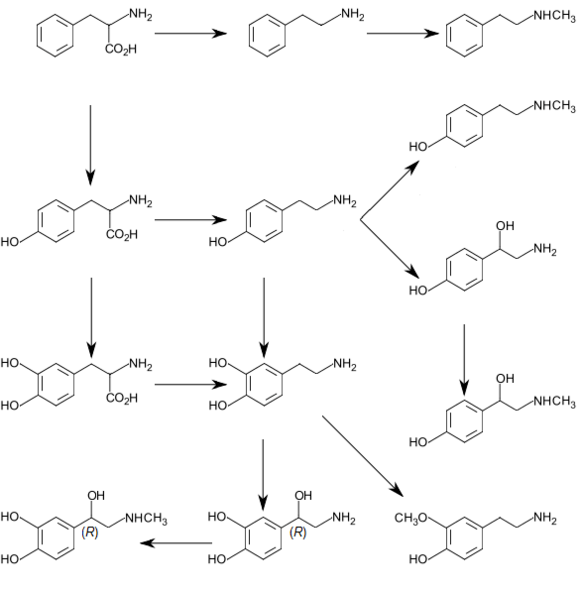
- Tyrosine (and its precursor phenylalanine) are precursors of the catecholamine neurotransmitters dopamine, epinephrine and norepinephrine and various trace amines.
- Phenylalanine is a precursor of phenethylamine and tyrosine in humans. In plants, it is a precursor of various phenylpropanoids, which are important in plant metabolism.
- Glycine is a precursor of porphyrins such as heme.[77]
- Arginine is a precursor of nitric oxide.[78]
- Ornithine and S-adenosylmethionine are precursors of polyamines.[79]
- Aspartate, glycine, and glutamine are precursors of nucleotides.[80] However, not all of the functions of other abundant non-standard amino acids are known.
Some non-standard amino acids are used as defenses against herbivores in plants.[81] For example, canavanine is an analogue of arginine that is found in many legumes,[82] and in particularly large amounts in Canavalia gladiata (sword bean).[83] This amino acid protects the plants from predators such as insects and can cause illness in people if some types of legumes are eaten without processing.[84] The non-protein amino acid mimosine is found in other species of legume, in particular Leucaena leucocephala.[85] This compound is an analogue of tyrosine and can poison animals that graze on these plants.
Uses in industry
Amino acids are used for a variety of applications in industry, but their main use is as additives to animal feed. This is necessary, since many of the bulk components of these feeds, such as soybeans, either have low levels or lack some of the essential amino acids: lysine, methionine, threonine, and tryptophan are most important in the production of these feeds.[86] In this industry, amino acids are also used to chelate metal cations in order to improve the absorption of minerals from supplements, which may be required to improve the health or production of these animals.[87]
The food industry is also a major consumer of amino acids, in particular, glutamic acid, which is used as a flavor enhancer,[88] and aspartame (aspartyl-phenylalanine-1-methyl ester) as a low-calorie artificial sweetener.[89] Similar technology to that used for animal nutrition is employed in the human nutrition industry to alleviate symptoms of mineral deficiencies, such as anemia, by improving mineral absorption and reducing negative side effects from inorganic mineral supplementation.[90]
The chelating ability of amino acids has been used in fertilizers for agriculture to facilitate the delivery of minerals to plants in order to correct mineral deficiencies, such as iron chlorosis. These fertilizers are also used to prevent deficiencies from occurring and improving the overall health of the plants.[91] The remaining production of amino acids is used in the synthesis of drugs and cosmetics.[86]
Similarly, some amino acids derivatives are used in pharmaceutical industry. They include 5-HTP (5-hydroxytryptophan) used for experimental treatment of depression,[92] L-DOPA (L-dihydroxyphenylalanine) for Parkinson's treatment,[93] and eflornithine drug that inhibits ornithine decarboxylase and used in the treatment of sleeping sickness.[94]
Expanded genetic code
Since 2001, 40 non-natural amino acids have been added into protein by creating a unique codon (recoding) and a corresponding transfer-RNA:aminoacyl – tRNA-synthetase pair to encode it with diverse physicochemical and biological properties in order to be used as a tool to exploring protein structure and function or to create novel or enhanced proteins.[16][17]
Nullomers
Nullomers are codons that in theory code for an amino acid, however in nature there is a selective bias against using this codon in favor of another, for example bacteria prefer to use CGA instead of AGA to code for arginine.[95] This creates some sequences that do not appear in the genome. This characteristic can be taken advantage of and used to create new selective cancer-fighting drugs[96] and to prevent cross-contamination of DNA samples from crime-scene investigations.[97]
Chemical building blocks
Amino acids are important as low-cost feedstocks. These compounds are used in chiral pool synthesis as enantiomerically pure building-blocks.[98]
Amino acids have been investigated as precursors chiral catalysts, e.g., for asymmetric hydrogenation reactions, although no commercial applications exist.[99]
Biodegradable plastics
Amino acids are under development as components of a range of biodegradable polymers. These materials have applications as environmentally friendly packaging and in medicine in drug delivery and the construction of prosthetic implants. These polymers include polypeptides, polyamides, polyesters, polysulfides, and polyurethanes with amino acids either forming part of their main chains or bonded as side chains. These modifications alter the physical properties and reactivities of the polymers.[100] An interesting example of such materials is polyaspartate, a water-soluble biodegradable polymer that may have applications in disposable diapers and agriculture.[101] Due to its solubility and ability to chelate metal ions, polyaspartate is also being used as a biodegradeable anti-scaling agent and a corrosion inhibitor.[102][103] In addition, the aromatic amino acid tyrosine is being developed as a possible replacement for toxic phenols such as bisphenol A in the manufacture of polycarbonates.[104]
Reactions
As amino acids have both a primary amine group and a primary carboxyl group, these chemicals can undergo most of the reactions associated with these functional groups. These include nucleophilic addition, amide bond formation, and imine formation for the amine group, and esterification, amide bond formation, and decarboxylation for the carboxylic acid group.[105] The combination of these functional groups allow amino acids to be effective polydentate ligands for metal-amino acid chelates.[106] The multiple side chains of amino acids can also undergo chemical reactions.[107] The types of these reactions are determined by the groups on these side chains and are, therefore, different between the various types of amino acid.
Chemical synthesis
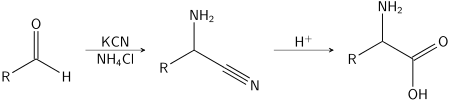
Several methods exist to synthesize amino acids. One of the oldest methods begins with the bromination at the α-carbon of a carboxylic acid. Nucleophilic substitution with ammonia then converts the alkyl bromide to the amino acid.[108] In alternative fashion, the Strecker amino acid synthesis involves the treatment of an aldehyde with potassium cyanide and ammonia, this produces an α-amino nitrile as an intermediate. Hydrolysis of the nitrile in acid then yields a α-amino acid.[109] Using ammonia or ammonium salts in this reaction gives unsubstituted amino acids, whereas substituting primary and secondary amines will yield substituted amino acids.[110] Likewise, using ketones, instead of aldehydes, gives α,α-disubstituted amino acids.[111] The classical synthesis gives racemic mixtures of α-amino acids as products, but several alternative procedures using asymmetric auxiliaries[112] or asymmetric catalysts[113][114] have been developed.[115]
At the current time, the most-adopted method is an automated synthesis on a solid support (e.g., polystyrene beads), using protecting groups (e.g., Fmoc and t-Boc) and activating groups (e.g., DCC and DIC).
Peptide bond formation
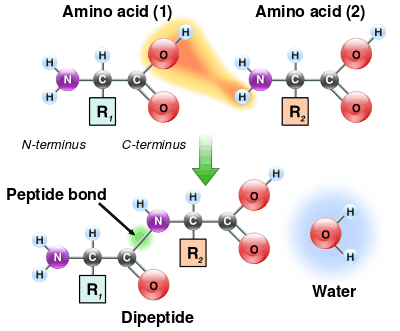
As both the amine and carboxylic acid groups of amino acids can react to form amide bonds, one amino acid molecule can react with another and become joined through an amide linkage. This polymerization of amino acids is what creates proteins. This condensation reaction yields the newly formed peptide bond and a molecule of water. In cells, this reaction does not occur directly; instead, the amino acid is first activated by attachment to a transfer RNA molecule through an ester bond. This aminoacyl-tRNA is produced in an ATP-dependent reaction carried out by an aminoacyl tRNA synthetase.[116] This aminoacyl-tRNA is then a substrate for the ribosome, which catalyzes the attack of the amino group of the elongating protein chain on the ester bond.[117] As a result of this mechanism, all proteins made by ribosomes are synthesized starting at their N-terminus and moving toward their C-terminus.
However, not all peptide bonds are formed in this way. In a few cases, peptides are synthesized by specific enzymes. For example, the tripeptide glutathione is an essential part of the defenses of cells against oxidative stress. This peptide is synthesized in two steps from free amino acids.[118] In the first step, gamma-glutamylcysteine synthetase condenses cysteine and glutamic acid through a peptide bond formed between the side chain carboxyl of the glutamate (the gamma carbon of this side chain) and the amino group of the cysteine. This dipeptide is then condensed with glycine by glutathione synthetase to form glutathione.[119]
In chemistry, peptides are synthesized by a variety of reactions. One of the most-used in solid-phase peptide synthesis uses the aromatic oxime derivatives of amino acids as activated units. These are added in sequence onto the growing peptide chain, which is attached to a solid resin support.[120] The ability to easily synthesize vast numbers of different peptides by varying the types and order of amino acids (using combinatorial chemistry) has made peptide synthesis particularly important in creating libraries of peptides for use in drug discovery through high-throughput screening.[121]
Biosynthesis
In plants, nitrogen is first assimilated into organic compounds in the form of glutamate, formed from alpha-ketoglutarate and ammonia in the mitochondrion. In order to form other amino acids, the plant uses transaminases to move the amino group to another alpha-keto carboxylic acid. For example, aspartate aminotransferase converts glutamate and oxaloacetate to alpha-ketoglutarate and aspartate.[122] Other organisms use transaminases for amino acid synthesis, too.
Nonstandard amino acids are usually formed through modifications to standard amino acids. For example, homocysteine is formed through the transsulfuration pathway or by the demethylation of methionine via the intermediate metabolite S-adenosyl methionine,[123] while hydroxyproline is made by a posttranslational modification of proline.[124]
Microorganisms and plants can synthesize many uncommon amino acids. For example, some microbes make 2-aminoisobutyric acid and lanthionine, which is a sulfide-bridged derivative of alanine. Both of these amino acids are found in peptidic lantibiotics such as alamethicin.[125] However, in plants, 1-aminocyclopropane-1-carboxylic acid is a small disubstituted cyclic amino acid that is a key intermediate in the production of the plant hormone ethylene.[126]
Catabolism

* Glucogenic, with the products having the ability to form glucose by gluconeogenesis
* Ketogenic, with the products not having the ability to form glucose. These products may still be used for ketogenesis or lipid synthesis.
* Amino acids catabolized into both glucogenic and ketogenic products.
Amino acids must first pass out of organelles and cells into blood circulation via amino acid transporters, since the amine and carboxylic acid groups are typically ionized. Degradation of an amino acid, occurring in the liver and kidneys, often involves deamination by moving its amino group to alpha-ketoglutarate, forming glutamate. This process involves transaminases, often the same as those used in amination during synthesis. In many vertebrates, the amino group is then removed through the urea cycle and is excreted in the form of urea. However, amino acid degradation can produce uric acid or ammonia instead. For example, serine dehydratase converts serine to pyruvate and ammonia.[80] After removal of one or more amino groups, the remainder of the molecule can sometimes be used to synthesize new amino acids, or it can be used for energy by entering glycolysis or the citric acid cycle, as detailed in image at right.
Physicochemical properties of amino acids
The 20 amino acids encoded directly by the genetic code can be divided into several groups based on their properties. Important factors are charge, hydrophilicity or hydrophobicity, size, and functional groups.[34] These properties are important for protein structure and protein–protein interactions. The water-soluble proteins tend to have their hydrophobic residues (Leu, Ile, Val, Phe, and Trp) buried in the middle of the protein, whereas hydrophilic side chains are exposed to the aqueous solvent. (Note that in biochemistry, a residue refers to a specific monomer within the polymeric chain of a polysaccharide, protein or nucleic acid.) The integral membrane proteins tend to have outer rings of exposed hydrophobic amino acids that anchor them into the lipid bilayer. In the case part-way between these two extremes, some peripheral membrane proteins have a patch of hydrophobic amino acids on their surface that locks onto the membrane. In similar fashion, proteins that have to bind to positively charged molecules have surfaces rich with negatively charged amino acids like glutamate and aspartate, while proteins binding to negatively charged molecules have surfaces rich with positively charged chains like lysine and arginine. There are different hydrophobicity scales of amino acid residues.[128]
Some amino acids have special properties such as cysteine, that can form covalent disulfide bonds to other cysteine residues, proline that forms a cycle to the polypeptide backbone, and glycine that is more flexible than other amino acids.
Many proteins undergo a range of posttranslational modifications, when additional chemical groups are attached to the amino acids in proteins. Some modifications can produce hydrophobic lipoproteins,[129] or hydrophilic glycoproteins.[130] These type of modification allow the reversible targeting of a protein to a membrane. For example, the addition and removal of the fatty acid palmitic acid to cysteine residues in some signaling proteins causes the proteins to attach and then detach from cell membranes.[131]
Table of standard amino acid abbreviations and properties
| Amino acid | 3-letter[132] | 1-letter[132] | Side chain
class |
Side chain
polarity[132] |
Side chain
charge (pH 7.4)[132] |
Hydropathy index[133] |
Absorbance
λmax(nm)[134] |
ε at
λmax (mM−1 cm−1)[134] |
MW
(weight) |
Occurrence
in proteins (%)[135] |
|---|---|---|---|---|---|---|---|---|---|---|
| Alanine | Ala | A | aliphatic | nonpolar | neutral | 1.8 | 89.094 | 8.76 | ||
| Arginine | Arg | R | basic | basic polar | positive | −4.5 | 174.203 | 5.78 | ||
| Asparagine | Asn | N | amide | polar | neutral | −3.5 | 132.119 | 3.93 | ||
| Aspartic acid | Asp | D | acid | acidic polar | negative | −3.5 | 133.104 | 5.49 | ||
| Cysteine | Cys | C | sulfur-containing | nonpolar | neutral | 2.5 | 250 | 0.3 | 121.154 | 1.38 |
| Glutamic acid | Glu | E | acid | acidic polar | negative | −3.5 | 147.131 | 6.32 | ||
| Glutamine | Gln | Q | amide | polar | neutral | −3.5 | 146.146 | 3.9 | ||
| Glycine | Gly | G | aliphatic | nonpolar | neutral | −0.4 | 75.067 | 7.03 | ||
| Histidine | His | H | basic aromatic | basic polar | positive(10%) neutral(90%) |
−3.2 | 211 | 5.9 | 155.156 | 2.26 |
| Isoleucine | Ile | I | aliphatic | nonpolar | neutral | 4.5 | 131.175 | 5.49 | ||
| Leucine | Leu | L | aliphatic | nonpolar | neutral | 3.8 | 131.175 | 9.68 | ||
| Lysine | Lys | K | basic | basic polar | positive | −3.9 | 146.189 | 5.19 | ||
| Methionine | Met | M | sulfur-containing | nonpolar | neutral | 1.9 | 149.208 | 2.32 | ||
| Phenylalanine | Phe | F | aromatic | nonpolar | neutral | 2.8 | 257, 206, 188 | 0.2, 9.3, 60.0 | 165.192 | 3.87 |
| Proline | Pro | P | cyclic | nonpolar | neutral | −1.6 | 115.132 | 5.02 | ||
| Serine | Ser | S | hydroxyl-containing | polar | neutral | −0.8 | 105.093 | 7.14 | ||
| Threonine | Thr | T | hydroxyl-containing | polar | neutral | −0.7 | 119.119 | 5.53 | ||
| Tryptophan | Trp | W | aromatic | nonpolar | neutral | −0.9 | 280, 219 | 5.6, 47.0 | 204.228 | 1.25 |
| Tyrosine | Tyr | Y | aromatic | polar | neutral | −1.3 | 274, 222, 193 | 1.4, 8.0, 48.0 | 181.191 | 2.91 |
| Valine | Val | V | aliphatic | nonpolar | neutral | 4.2 | 117.148 | 6.73 |
Two additional amino acids are in some species coded for by codons that are usually interpreted as stop codons:
| 21st and 22nd amino acids | 3-letter | 1-letter | MW (weight) |
|---|---|---|---|
| Selenocysteine | Sec | U | 168.064 |
| Pyrrolysine | Pyl | O | 255.313 |
In addition to the specific amino acid codes, placeholders are used in cases where chemical or crystallographic analysis of a peptide or protein cannot conclusively determine the identity of a residue. They are also used to summarise conserved protein sequence motifs. The use of single letters to indicate sets of similar residues is similar to the use of abbreviation codes for degenerate bases.[136][137]
| Ambiguous amino acids | 3-letter | 1-letter | |
|---|---|---|---|
| Any / unknown | Xaa | X | All |
| Asparagine or aspartic acid | Asx | B | D, N |
| Glutamine or glutamic acid | Glx | Z | E, Q |
| Leucine or Isoleucine | Xle | J | I, L |
| Hydrophobic | Φ | V, I, L, F, W, Y, M | |
| Aromatic | Ω | F, W, Y, H | |
| Aliphatic (non-aromatic) | Ψ | V, I, L, M | |
| Small | π | P, G, A, S | |
| Hydrophilic | ζ | S, T, H, N, Q, E, D, K, R | |
| Positively charged | + | K, R, H | |
| Negatively charged | − | D, E |
Unk is sometimes used instead of Xaa, but is less standard.
In addition, many non-standard amino acids have a specific code. For example, several peptide drugs, such as Bortezomib and MG132, are artificially synthesized and retain their protecting groups, which have specific codes. Bortezomib is Pyz-Phe-boroLeu, and MG132 is Z-Leu-Leu-Leu-al. To aid in the analysis of protein structure, photo-reactive amino acid analogs are available. These include photoleucine (pLeu) and photomethionine (pMet).[138]
See also
- Amino acid dating
- Beta-peptide
- Degron
- Erepsin
- Homochirality
- Hyperaminoacidemia
- Leucines
- Miller–Urey experiment
- Nucleic acid sequence
- Proteinogenic amino acid
- Table of codons, 3-nucleotide sequences that encode each amino acid
References and notes
- ↑ "Amino". Dictionary.com. 2015. Retrieved 3 July 2015.
- ↑ "amino acid". Cambridge Dictionaries Online. Cambridge University Press. 2015. Retrieved 3 July 2015.
- ↑ "amino". FreeDictionary.com. Farlex. 2015. Retrieved 3 July 2015.
- ↑ Wagner I, Musso H (November 1983). "New Naturally Occurring Amino Acids". Angewandte Chemie International Edition in English. 22 (11): 816–28. doi:10.1002/anie.198308161.

- ↑ Latham MC (1997). "Chapter 8. Body composition, the functions of food, metabolism and energy". Human nutrition in the developing world. Food and Nutrition Series – No. 29. Rome: Food and Agriculture Organization of the United Nations.
- ↑ Proline is an exception to this general formula. It lacks the NH2 group because of the cyclization of the side chain and is known as an imino acid; it falls under the category of special structured amino acids.
- ↑ Clark, Jim (August 2007). "an introduction to amino acids". chemguide. Retrieved 4 July 2015.
- ↑ Jakubke H, Sewald N (2008). "Amino acids". Peptides from A to Z: A Concise Encyclopedia. Germany: Wiley-VCH. p. 20. ISBN 9783527621170 – via Google Books.
- ↑ Pollegioni L, Servi S, eds. (2012). Unnatural Amino Acids: Methods and Protocols. Methods in Molecular Biology – Volume 794. Humana Press. p. v. ISBN 978-1-61779-331-8. OCLC 756512314.
- ↑ Hertweck C (October 2011). "Biosynthesis and Charging of Pyrrolysine, the 22nd Genetically Encoded Amino Acid". Angewandte Chemie International Edition. 50 (41): 9540–1. PMID 21796749. doi:10.1002/anie.201103769.
- ↑ "Chapter 1: Proteins are the Body's Worker Molecules". The Structures of Life. National Institute of General Medical Sciences. 27 October 2011. Retrieved 20 May 2008.
- ↑ Michal G, Schomburg D, eds. (2012). Biochemical Pathways: An Atlas of Biochemistry and Molecular Biology (2nd ed.). Oxford: Wiley-Blackwell. p. 5. ISBN 978-0-470-14684-2.
- 1 2 Modeling Electrostatic Contributions to Protein Folding and Binding – Tjong, p.1 footnote
- 1 2 Frontiers in Drug Design and Discovery ed. Atta-Ur-Rahman & others, p.299
- 1 2 Elzanowski A, Ostell J (7 April 2008). "The Genetic Codes". National Center for Biotechnology Information (NCBI). Retrieved 10 March 2010.
- 1 2 Xie J, Schultz PG (December 2005). "Adding amino acids to the genetic repertoire". Current Opinion in Chemical Biology. 9 (6): 548–54. PMID 16260173. doi:10.1016/j.cbpa.2005.10.011.
- 1 2 Wang Q, Parrish AR, Wang L (March 2009). "Expanding the genetic code for biological studies". Chemistry & Biology. 16 (3): 323–36. PMC 2696486
 . PMID 19318213. doi:10.1016/j.chembiol.2009.03.001.
. PMID 19318213. doi:10.1016/j.chembiol.2009.03.001. - ↑ Simon M (2005). Emergent computation: emphasizing bioinformatics. New York: AIP Press/Springer Science+Business Media. pp. 105–106. ISBN 0-387-22046-1.
- ↑ Petroff OA (December 2002). "GABA and glutamate in the human brain". The Neuroscientist. 8 (6): 562–73. PMID 12467378. doi:10.1177/1073858402238515.
- ↑ For example, ruminants such as cows obtain a number of amino acids via microbes in the first two stomach chambers.
- ↑ Vauquelin LN, Robiquet PJ (1806). "The discovery of a new plant principle in Asparagus sativus". Annales de Chimie. 57: 88–93.
- 1 2 Anfinsen CB, Edsall JT, Richards FM (1972). Advances in Protein Chemistry. New York: Academic Press. pp. 99, 103. ISBN 978-0-12-034226-6.
- ↑ Wollaston WH (1810). "On cystic oxide, a new species of urinary calculus". Philosophical Transactions of the Royal Society. 100: 223–30. doi:10.1098/rstl.1810.0015.
- ↑ Baumann E (1884). "Über cystin und cystein". Z Physiol Chem. 8 (4): 299–305. Archived from the original on 14 March 2011. Retrieved 28 March 2011.
- ↑ Braconnot HM (1820). "Sur la conversion des matières animales en nouvelles substances par le moyen de l'acide sulfurique". Annales de Chimie et de Physique. 2nd Series. 13: 113–25.
- ↑ Simoni RD, Hill RL, Vaughan M (September 2002). "The discovery of the amino acid threonine: the work of William C. Rose [classical article]". The Journal of Biological Chemistry. 277 (37): E25. PMID 12218068.
- ↑ McCoy RH, Meyer CE, Rose WC (1935). "Feeding Experiments with Mixtures of Highly Purified Amino Acids. VIII. Isolation and Identification of a New Essential Amino Acid". Journal of Biological Chemistry. 112: 283–302.
- ↑ "etymonline.com entry for amino". www.etymonline.com. Retrieved 19 July 2010.
- ↑ Fruton JS (1990). "Chapter 5- Emil Fischer and Franz Hofmeister". Contrasts in Scientific Style: Research Groups in the Chemical and Biochemical Sciences,. 191. American Philosophical Society. pp. 163–165. ISBN 0-87169-191-4.
- ↑ "Alpha amino acid – Medical definition". Merriam-Webster dictionary.
- ↑ Proline at the US National Library of Medicine Medical Subject Headings (MeSH)
- ↑ "Archived copy". Archived from the original on 18 January 2008. Retrieved 3 January 2015.
- ↑ IUPAC, Compendium of Chemical Terminology, 2nd ed. (the "Gold Book") (1997). Online corrected version: (2006–) "Imino acids".
- 1 2 3 4 5 6 Creighton TH (1993). "Chapter 1". Proteins: structures and molecular properties. San Francisco: W. H. Freeman. ISBN 978-0-7167-7030-5.
- ↑ Pisarewicz K, Mora D, Pflueger FC, Fields GB, Marí F (May 2005). "Polypeptide chains containing D-gamma-hydroxyvaline". Journal of the American Chemical Society. 127 (17): 6207–15. PMID 15853325. doi:10.1021/ja050088m.
- ↑ van Heijenoort J (March 2001). "Formation of the glycan chains in the synthesis of bacterial peptidoglycan". Glycobiology. 11 (3): 25R–36R. PMID 11320055. doi:10.1093/glycob/11.3.25R.
- ↑ Wolosker H, Dumin E, Balan L, Foltyn VN (July 2008). "D-amino acids in the brain: D-serine in neurotransmission and neurodegeneration". The FEBS Journal. 275 (14): 3514–26. PMID 18564180. doi:10.1111/j.1742-4658.2008.06515.x.
- ↑ Matthews BW (June 2009). "Racemic crystallography—easy crystals and easy structures: What's not to like?". Protein Science. 18 (6): 1135–8. PMC 2774423 . PMID 19472321. doi:10.1002/pro.125.
- ↑ Hatem, Salama Mohamed Ali (2006). "Gas chromatographic determination of Amino Acid Enantiomers in tobacco and bottled wines". University of Giessen. Retrieved 17 November 2008.
- ↑ "Nomenclature and Symbolism for Amino Acids and Peptides". IUPAC-IUB Joint Commission on Biochemical Nomenclature. 1983. Archived from the original on 9 October 2008. Retrieved 17 November 2008.
- ↑ Jodidi, S. L. (1 March 1926). "The Formol Titration of Certain Amino Acids". Journal of the American Chemical Society. 48 (3): 751–753. doi:10.1021/ja01414a033.
- ↑ Liebecq C, ed. (1992). Biochemical Nomenclature and Related Documents (2nd ed.). Portland Press. pp. 39–69. ISBN 978-1-85578-005-7.
- ↑ Smith AD (1997). Oxford dictionary of biochemistry and molecular biology. Oxford: Oxford University Press. p. 535. ISBN 978-0-19-854768-6. OCLC 37616711.
- ↑ Simmons WJ, Meisenberg G (2006). Principles of medical biochemistry. Mosby Elsevier. p. 19. ISBN 0-323-02942-6.
- ↑ Fennema OR. Food Chemistry 3rd Ed. CRC Press. pp. 327–8. ISBN 0-8247-9691-8.
- ↑ Rodnina MV, Beringer M, Wintermeyer W (January 2007). "How ribosomes make peptide bonds". Trends in Biochemical Sciences. 32 (1): 20–6. PMID 17157507. doi:10.1016/j.tibs.2006.11.007.
- ↑ Driscoll DM, Copeland PR (2003). "Mechanism and regulation of selenoprotein synthesis". Annual Review of Nutrition. 23 (1): 17–40. PMID 12524431. doi:10.1146/annurev.nutr.23.011702.073318.
- ↑ Krzycki JA (December 2005). "The direct genetic encoding of pyrrolysine". Current Opinion in Microbiology. 8 (6): 706–12. PMID 16256420. doi:10.1016/j.mib.2005.10.009.
- ↑ Théobald-Dietrich A, Giegé R, Rudinger-Thirion J (2005). "Evidence for the existence in mRNAs of a hairpin element responsible for ribosome dependent pyrrolysine insertion into proteins". Biochimie. 87 (9–10): 813–7. PMID 16164991. doi:10.1016/j.biochi.2005.03.006.
- ↑ Vermeer C (March 1990). "Gamma-carboxyglutamate-containing proteins and the vitamin K-dependent carboxylase". The Biochemical Journal. 266 (3): 625–36. PMC 1131186 . PMID 2183788. doi:10.1042/bj2660625.
- ↑ Bhattacharjee A, Bansal M (March 2005). "Collagen structure: the Madras triple helix and the current scenario". IUBMB Life. 57 (3): 161–72. PMID 16036578. doi:10.1080/15216540500090710.
- ↑ Park MH (February 2006). "The post-translational synthesis of a polyamine-derived amino acid, hypusine, in the eukaryotic translation initiation factor 5A (eIF5A)". Journal of Biochemistry. 139 (2): 161–9. PMC 2494880 . PMID 16452303. doi:10.1093/jb/mvj034.
- ↑ Blenis J, Resh MD (December 1993). "Subcellular localization specified by protein acylation and phosphorylation". Current Opinion in Cell Biology. 5 (6): 984–9. PMID 8129952. doi:10.1016/0955-0674(93)90081-Z.
- ↑ Curis E, Nicolis I, Moinard C, Osowska S, Zerrouk N, Bénazeth S, Cynober L (November 2005). "Almost all about citrulline in mammals". Amino Acids. 29 (3): 177–205. PMID 16082501. doi:10.1007/s00726-005-0235-4.
- ↑ Coxon KM, Chakauya E, Ottenhof HH, Whitney HM, Blundell TL, Abell C, Smith AG (August 2005). "Pantothenate biosynthesis in higher plants". Biochemical Society Transactions. 33 (Pt 4): 743–6. PMID 16042590. doi:10.1042/BST0330743.
- ↑ Ketchem RR, Hu W, Cross TA (September 1993). "High-resolution conformation of gramicidin A in a lipid bilayer by solid-state NMR". Science. 261 (5127): 1457–60. PMID 7690158. doi:10.1126/science.7690158.
- ↑ Khoury GA, Baliban RC, Floudas CA (September 2011). "Proteome-wide post-translational modification statistics: frequency analysis and curation of the swiss-prot database". Scientific Reports. 1 (90). PMC 3201773 . PMID 22034591. doi:10.1038/srep00090.
- ↑ Kryukov GV, Castellano S, Novoselov SV, Lobanov AV, Zehtab O, Guigó R, Gladyshev VN (May 2003). "Characterization of mammalian selenoproteomes". Science. 300 (5624): 1439–43. Bibcode:2003Sci...300.1439K. PMID 12775843. doi:10.1126/science.1083516.
- ↑ Gromer S, Urig S, Becker K (January 2004). "The thioredoxin system—from science to clinic". Medicinal Research Reviews. 24 (1): 40–89. PMID 14595672. doi:10.1002/med.10051.
- ↑ Sakami W, Harrington H (1963). "Amino acid metabolism". Annual Review of Biochemistry. 32 (1): 355–98. PMID 14144484. doi:10.1146/annurev.bi.32.070163.002035.
- ↑ Brosnan JT (April 2000). "Glutamate, at the interface between amino acid and carbohydrate metabolism". The Journal of Nutrition. 130 (4S Suppl): 988S–90S. PMID 10736367.
- ↑ Young VR, Ajami AM (September 2001). "Glutamine: the emperor or his clothes?". The Journal of Nutrition. 131 (9 Suppl): 2449S–59S; discussion 2486S–7S. PMID 11533293.
- ↑ Young VR (August 1994). "Adult amino acid requirements: the case for a major revision in current recommendations". The Journal of Nutrition. 124 (8 Suppl): 1517S–1523S. PMID 8064412.
- ↑ Fürst P, Stehle P (June 2004). "What are the essential elements needed for the determination of amino acid requirements in humans?". The Journal of Nutrition. 134 (6 Suppl): 1558S–1565S. PMID 15173430.
- ↑ Reeds PJ (July 2000). "Dispensable and indispensable amino acids for humans". The Journal of Nutrition. 130 (7): 1835S–40S. PMID 10867060.
- ↑ Imura K, Okada A (January 1998). "Amino acid metabolism in pediatric patients". Nutrition. 14 (1): 143–8. PMID 9437700. doi:10.1016/S0899-9007(97)00230-X.
- ↑ Lourenço R, Camilo ME (2002). "Taurine: a conditionally essential amino acid in humans? An overview in health and disease". Nutricion Hospitalaria. 17 (6): 262–70. PMID 12514918.
- ↑ Holtcamp W (March 2012). "The emerging science of BMAA: do cyanobacteria contribute to neurodegenerative disease?". Environmental Health Perspectives. 120 (3): A110–6. PMC 3295368 . PMID 22382274. doi:10.1289/ehp.120-a110.
- ↑ Cox PA, Davis DA, Mash DC, Metcalf JS, Banack SA (January 2016). "Dietary exposure to an environmental toxin triggers neurofibrillary tangles and amyloid deposits in the brain". Proceedings. Biological Sciences. 283 (1823): 20152397. PMC 4795023 . PMID 26791617. doi:10.1098/rspb.2015.2397.
- 1 2 Brook MS, Wilkinson DJ, Phillips BE, Perez-Schindler J, Philp A, Smith K, Atherton PJ (January 2016). "Skeletal muscle homeostasis and plasticity in youth and ageing: impact of nutrition and exercise". Acta Physiologica. 216 (1): 15–41. PMC 4843955 . PMID 26010896. doi:10.1111/apha.12532.
- ↑ Lipton JO, Sahin M (October 2014). "The neurology of mTOR". Neuron. 84 (2): 275–91. PMC 4223653 . PMID 25374355. doi:10.1016/j.neuron.2014.09.034.
Figure 2: The mTOR Signaling Pathway - 1 2 Phillips SM (May 2014). "A brief review of critical processes in exercise-induced muscular hypertrophy". Sports Medicine. 44 Suppl 1: S71–7. PMC 4008813 . PMID 24791918. doi:10.1007/s40279-014-0152-3.
- ↑ Broadley KJ (March 2010). "The vascular effects of trace amines and amphetamines". Pharmacol. Ther. 125 (3): 363–375. PMID 19948186. doi:10.1016/j.pharmthera.2009.11.005.
- ↑ Lindemann L, Hoener MC (May 2005). "A renaissance in trace amines inspired by a novel GPCR family". Trends Pharmacol. Sci. 26 (5): 274–281. PMID 15860375. doi:10.1016/j.tips.2005.03.007.
- ↑ Wang X, Li J, Dong G, Yue J (February 2014). "The endogenous substrates of brain CYP2D". Eur. J. Pharmacol. 724: 211–218. PMID 24374199. doi:10.1016/j.ejphar.2013.12.025.
The highest level of brain CYP2D activity was found in the substantia nigra ... The in vitro and in vivo studies have shown the contribution of the alternative CYP2D-mediated dopamine synthesis to the concentration of this neurotransmitter although the classic biosynthetic route to dopamine from tyrosine is active. ... Tyramine levels are especially high in the basal ganglia and limbic system, which are thought to be related to individual behavior and emotion (Yu et al., 2003c). ... Rat CYP2D isoforms (2D2/2D4/2D18) are less efficient than human CYP2D6 for the generation of dopamine from p-tyramine. The Km values of the CYP2D isoforms are as follows: CYP2D6 (87–121 μm) ≈ CYP2D2 ≈ CYP2D18 > CYP2D4 (256 μm) for m-tyramine and CYP2D4 (433 μm) > CYP2D2 ≈ CYP2D6 > CYP2D18 (688 μm) for p-tyramine
- ↑ Savelieva KV, Zhao S, Pogorelov VM, Rajan I, Yang Q, Cullinan E, Lanthorn TH (2008). Bartolomucci A, ed. "Genetic disruption of both tryptophan hydroxylase genes dramatically reduces serotonin and affects behavior in models sensitive to antidepressants". PloS One. 3 (10): e3301. Bibcode:2008PLoSO...3.3301S. PMC 2565062 . PMID 18923670. doi:10.1371/journal.pone.0003301.
- ↑ Shemin D, Rittenberg D (December 1946). "The biological utilization of glycine for the synthesis of the protoporphyrin of hemoglobin". The Journal of Biological Chemistry. 166 (2): 621–5. PMID 20276176.
- ↑ Tejero J, Biswas A, Wang ZQ, Page RC, Haque MM, Hemann C, Zweier JL, Misra S, Stuehr DJ (November 2008). "Stabilization and characterization of a heme-oxy reaction intermediate in inducible nitric-oxide synthase". The Journal of Biological Chemistry. 283 (48): 33498–507. PMC 2586280 . PMID 18815130. doi:10.1074/jbc.M806122200.
- ↑ Rodríguez-Caso C, Montañez R, Cascante M, Sánchez-Jiménez F, Medina MA (August 2006). "Mathematical modeling of polyamine metabolism in mammals". The Journal of Biological Chemistry. 281 (31): 21799–812. PMID 16709566. doi:10.1074/jbc.M602756200.
- 1 2 Stryer L, Berg JM, Tymoczko JL (2002). Biochemistry (5th ed.). New York: W.H. Freeman. pp. 693–8. ISBN 978-0-7167-4684-3.
- ↑ Hylin, John W. (1969). "Toxic peptides and amino acids in foods and feeds". Journal of Agricultural and Food Chemistry. 17 (3): 492–6. doi:10.1021/jf60163a003.
- ↑ Turner BL, Harborne JB (1967). "Distribution of canavanine in the plant kingdom". Phytochemistry. 6 (6): 863–66. doi:10.1016/S0031-9422(00)86033-1.
- ↑ Ekanayake S, Skog K, Asp NG (May 2007). "Canavanine content in sword beans (Canavalia gladiata): analysis and effect of processing". Food and Chemical Toxicology. 45 (5): 797–803. PMID 17187914. doi:10.1016/j.fct.2006.10.030.
- ↑ Rosenthal GA (2001). "L-Canavanine: a higher plant insecticidal allelochemical". Amino Acids. 21 (3): 319–30. PMID 11764412. doi:10.1007/s007260170017.
- ↑ Hammond AC (May 1995). "Leucaena toxicosis and its control in ruminants". Journal of Animal Science. 73 (5): 1487–92. PMID 7665380.
- 1 2 Leuchtenberger W, Huthmacher K, Drauz K (November 2005). "Biotechnological production of amino acids and derivatives: current status and prospects". Applied Microbiology and Biotechnology. 69 (1): 1–8. PMID 16195792. doi:10.1007/s00253-005-0155-y.
- ↑ Ashmead HD (1993). The Role of Amino Acid Chelates in Animal Nutrition. Westwood: Noyes Publications.
- ↑ Garattini S (April 2000). "Glutamic acid, twenty years later". The Journal of Nutrition. 130 (4S Suppl): 901S–9S. PMID 10736350.
- ↑ Stegink LD (July 1987). "The aspartame story: a model for the clinical testing of a food additive". The American Journal of Clinical Nutrition. 46 (1 Suppl): 204–15. PMID 3300262.
- ↑ Albion Laboratories, Inc. "Albion Ferrochel Website". Retrieved 12 July 2011.
- ↑ Ashmead HD (1986). Foliar Feeding of Plants with Amino Acid Chelates. Park Ridge: Noyes Publications.
- ↑ Turner EH, Loftis JM, Blackwell AD (March 2006). "Serotonin a la carte: supplementation with the serotonin precursor 5-hydroxytryptophan". Pharmacology & Therapeutics. 109 (3): 325–38. PMID 16023217. doi:10.1016/j.pharmthera.2005.06.004.
- ↑ Kostrzewa RM, Nowak P, Kostrzewa JP, Kostrzewa RA, Brus R (March 2005). "Peculiarities of L: -DOPA treatment of Parkinson's disease". Amino Acids. 28 (2): 157–64. PMID 15750845. doi:10.1007/s00726-005-0162-4.
- ↑ Heby O, Persson L, Rentala M (August 2007). "Targeting the polyamine biosynthetic enzymes: a promising approach to therapy of African sleeping sickness, Chagas' disease, and leishmaniasis". Amino Acids. 33 (2): 359–66. PMID 17610127. doi:10.1007/s00726-007-0537-9.
- ↑ Cruz-Vera LR, Magos-Castro MA, Zamora-Romo E, Guarneros G (2004). "Ribosome stalling and peptidyl-tRNA drop-off during translational delay at AGA codons". Nucleic Acids Research. 32 (15): 4462–8. PMC 516057 . PMID 15317870. doi:10.1093/nar/gkh784.
- ↑ Andy C (October 2012). "Molecules 'too dangerous for nature' kill cancer cells". New Scientist.
- ↑ "Lethal DNA tags could keep innocent people out of jail". New Scientist. 2 May 2013.
- ↑ Hanessian S (1993). "Reflections on the total synthesis of natural products: Art, craft, logic, and the chiron approach". Pure and Applied Chemistry. 65 (6): 1189–204. doi:10.1351/pac199365061189.
- ↑ Blaser HU (1992). "The chiral pool as a source of enantioselective catalysts and auxiliaries". Chemical Reviews. 92 (5): 935–52. doi:10.1021/cr00013a009.
- ↑ Sanda F, Endo T (1999). "Syntheses and functions of polymers based on amino acids". Macromolecular Chemistry and Physics. 200 (12): 2651–61. doi:10.1002/(SICI)1521-3935(19991201)200:12<2651::AID-MACP2651>3.0.CO;2-P.
- ↑ Gross RA, Kalra B (August 2002). "Biodegradable polymers for the environment". Science. 297 (5582): 803–7. Bibcode:2002Sci...297..803G. PMID 12161646. doi:10.1126/science.297.5582.803.
- ↑ Low KC, Wheeler AP, Koskan LP (1996). Commercial poly(aspartic acid) and Its Uses. Advances in Chemistry Series. 248. Washington, D.C.: American Chemical Society.
- ↑ Thombre SM, Sarwade BD (2005). "Synthesis and Biodegradability of Polyaspartic Acid: A Critical Review". Journal of Macromolecular Science, Part A. 42 (9): 1299–1315. doi:10.1080/10601320500189604.
- ↑ Bourke SL, Kohn J (April 2003). "Polymers derived from the amino acid L-tyrosine: polycarbonates, polyarylates and copolymers with poly(ethylene glycol)". Advanced Drug Delivery Reviews. 55 (4): 447–66. PMID 12706045. doi:10.1016/S0169-409X(03)00038-3.
- ↑ Elmore DT, Barrett GC (1998). Amino acids and peptides. Cambridge, UK: Cambridge University Press. pp. 48–60. ISBN 0-521-46827-2.
- ↑ Konara S, Gagnona K, Clearfield A, Thompson C, Hartle J, Ericson C, Nelson C (2010). "Structural determination and characterization of copper and zinc bis-glycinates with X-ray crystallography and mass spectrometry". Journal of Coordination Chemistry. 63 (19): 3335–47. doi:10.1080/00958972.2010.514336.
- ↑ Gutteridge A, Thornton JM (November 2005). "Understanding nature's catalytic toolkit". Trends in Biochemical Sciences. 30 (11): 622–9. PMID 16214343. doi:10.1016/j.tibs.2005.09.006.
- ↑ McMurry J (1996). Organic chemistry. Pacific Grove, CA, USA: Brooks/Cole. p. 1064. ISBN 0-534-23832-7.
- ↑ Strecker A (1850). "Ueber die künstliche Bildung der Milchsäure und einen neuen, dem Glycocoll homologen Körper". Justus Liebigs Annalen der Chemie. 75 (1): 27–45. doi:10.1002/jlac.18500750103.
- ↑ Strecker A (1854). "Ueber einen neuen aus Aldehyd – Ammoniak und Blausäure entstehenden Körper". Justus Liebigs Annalen der Chemie. 91 (3): 349–51. doi:10.1002/jlac.18540910309.
- ↑ Masumoto S, Usuda H, Suzuki M, Kanai M, Shibasaki M (May 2003). "Catalytic enantioselective Strecker reaction of ketoimines". Journal of the American Chemical Society. 125 (19): 5634–5. PMID 12733893. doi:10.1021/ja034980.
- ↑ Davis FA, Reddy RE, Portonovo PS (1994). "Asymmetric strecker synthesis using enantiopure sulfinimines: A convenient synthesis of α-amino acids". Tetrahedron Letters. 35 (50): 9351–9354. doi:10.1016/S0040-4039(00)78540-6.
- ↑ Ishitani H, Komiyama S, Hasegawa Y, Kobayashi S (2000). "Catalytic Asymmetric Strecker Synthesis. Preparation of Enantiomerically Pure α-Amino Acid Derivatives from Aldimines and Tributyltin Cyanide or Achiral Aldehydes, Amines, and Hydrogen Cyanide Using a Chiral Zirconium Catalyst". Journal of the American Chemical Society. 122 (5): 762–6. doi:10.1021/ja9935207.
- ↑ Huang J, Corey EJ (December 2004). "A new chiral catalyst for the enantioselective Strecker synthesis of alpha-amino acids". Organic Letters. 6 (26): 5027–9. PMID 15606127. doi:10.1021/ol047698w.
- ↑ Duthaler RO (1994). "Recent developments in the stereoselective synthesis of α-aminoacids". Tetrahedron. 50 (6): 1539–1650. doi:10.1016/S0040-4020(01)80840-1.
- ↑ Ibba M, Söll D (May 2001). "The renaissance of aminoacyl-tRNA synthesis". EMBO Reports. 2 (5): 382–7. PMC 1083889 . PMID 11375928. doi:10.1093/embo-reports/kve095.
- ↑ Lengyel P, Söll D (June 1969). "Mechanism of protein biosynthesis". Bacteriological Reviews. 33 (2): 264–301. PMC 378322 . PMID 4896351.
- ↑ Wu G, Fang YZ, Yang S, Lupton JR, Turner ND (March 2004). "Glutathione metabolism and its implications for health". The Journal of Nutrition. 134 (3): 489–92. PMID 14988435.
- ↑ Meister A (November 1988). "Glutathione metabolism and its selective modification". The Journal of Biological Chemistry. 263 (33): 17205–8. PMID 3053703.
- ↑ Carpino LA (1992). "1-Hydroxy-7-azabenzotriazole. An efficient peptide coupling additive". Journal of the American Chemical Society. 115 (10): 4397–8. doi:10.1021/ja00063a082.
- ↑ Marasco D, Perretta G, Sabatella M, Ruvo M (October 2008). "Past and future perspectives of synthetic peptide libraries". Current Protein & Peptide Science. 9 (5): 447–67. PMID 18855697. doi:10.2174/138920308785915209.
- ↑ Jones RC, Buchanan BB, Gruissem W (2000). Biochemistry & molecular biology of plants. Rockville, Md: American Society of Plant Physiologists. pp. 371–2. ISBN 0-943088-39-9.
- ↑ Brosnan JT, Brosnan ME (June 2006). "The sulfur-containing amino acids: an overview". The Journal of Nutrition. 136 (6 Suppl): 1636S–1640S. PMID 16702333.
- ↑ Kivirikko KI, Pihlajaniemi T (1998). "Collagen hydroxylases and the protein disulfide isomerase subunit of prolyl 4-hydroxylases". Advances in Enzymology and Related Areas of Molecular Biology. Advances in Enzymology – and Related Areas of Molecular Biology. 72: 325–98. ISBN 9780470123188. PMID 9559057. doi:10.1002/9780470123188.ch9.
- ↑ Whitmore L, Wallace BA (May 2004). "Analysis of peptaibol sequence composition: implications for in vivo synthesis and channel formation". European Biophysics Journal. 33 (3): 233–7. PMID 14534753. doi:10.1007/s00249-003-0348-1.
- ↑ Alexander L, Grierson D (October 2002). "Ethylene biosynthesis and action in tomato: a model for climacteric fruit ripening". Journal of Experimental Botany. 53 (377): 2039–55. PMID 12324528. doi:10.1093/jxb/erf072.
- ↑ Stipanuk, M. H. (2006). Biochemical, physiological, & molecular aspects of human nutrition (2 ed.): Saunders Elsevier.
- ↑ Urry DW (2004). "The change in Gibbs free energy for hydrophobic association: Derivation and evaluation by means of inverse temperature transitions". Chemical Physics Letters. 399 (1–3): 177–83. doi:10.1016/S0009-2614(04)01565-9.
- ↑ Magee T, Seabra MC (April 2005). "Fatty acylation and prenylation of proteins: what's hot in fat". Current Opinion in Cell Biology. 17 (2): 190–6. PMID 15780596. doi:10.1016/j.ceb.2005.02.003.
- ↑ Pilobello KT, Mahal LK (June 2007). "Deciphering the glycocode: the complexity and analytical challenge of glycomics". Current Opinion in Chemical Biology. 11 (3): 300–5. PMID 17500024. doi:10.1016/j.cbpa.2007.05.002.
- ↑ Smotrys JE, Linder ME (2004). "Palmitoylation of intracellular signaling proteins: regulation and function". Annual Review of Biochemistry. 73 (1): 559–87. PMID 15189153. doi:10.1146/annurev.biochem.73.011303.073954.
- 1 2 3 4 Hausman, Robert E., Cooper, Geoffrey M. (2004). The cell: a molecular approach. Washington, D.C: ASM Press. p. 51. ISBN 0-87893-214-3.
- ↑ Kyte J, Doolittle RF (May 1982). "A simple method for displaying the hydropathic character of a protein". Journal of Molecular Biology. 157 (1): 105–32. PMID 7108955. doi:10.1016/0022-2836(82)90515-0.
- 1 2 Freifelder D (1983). Physical Biochemistry (2nd ed.). W. H. Freeman and Company. ISBN 0-7167-1315-2.
- ↑ Kozlowski LP (January 2017). "Proteome-pI: proteome isoelectric point database". Nucleic Acids Research. 45 (D1): D1112–D1116. PMC 5210655 . PMID 27789699. doi:10.1093/nar/gkw978.
- ↑ Aasland R, Abrams C, Ampe C, Ball LJ, Bedford MT, Cesareni G, Gimona M, Hurley JH, Jarchau T, Lehto VP, Lemmon MA, Linding R, Mayer BJ, Nagai M, Sudol M, Walter U, Winder SJ (February 2002). "Normalization of nomenclature for peptide motifs as ligands of modular protein domains". FEBS Letters. 513 (1): 141–4. PMID 11911894. doi:10.1111/j.1432-1033.1968.tb00350.x.
- ↑ IUPAC-IUB Commission on Biochemical Nomenclature (1972). "A one-letter notation for amino acid sequences". Pure and Applied Chemistry. Chimie Pure et Appliquee. 31 (4): 641–5. PMID 5080161.
- ↑ Suchanek M, Radzikowska A, Thiele C (April 2005). "Photo-leucine and photo-methionine allow identification of protein-protein interactions in living cells". Nature Methods. 2 (4): 261–7. PMID 15782218. doi:10.1038/nmeth752.
Further reading
- Tymoczko JL (2012). "Protein Composition and Structure". Biochemistry. New York: W. H. Freeman and company. pp. 28–31. ISBN 9781429229364.
- Doolittle RF (1989). "Redundancies in protein sequences". In Fasman, G.D. Predictions of Protein Structure and the Principles of Protein Conformation. New York: Plenum Press. pp. 599–623. ISBN 978-0-306-43131-9. LCCN 89008555.
- Nelson DL, Cox MM (2000). Lehninger Principles of Biochemistry (3rd ed.). Worth Publishers. ISBN 978-1-57259-153-0. LCCN 99049137.
- Meierhenrich U (2008). Amino acids and the asymmetry of life (PDF). Berlin: Springer Verlag. ISBN 978-3-540-76885-2. LCCN 2008930865. Archived from the original on 12 January 2012.
External links
 Media related to Amino acid at Wikimedia Commons
Media related to Amino acid at Wikimedia Commons
| General topics |  | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| By properties |
| ||||||||||
| Other classifications | |||||||||||