Coelophysis bauri
| Coelophysis bauri Temporal range: Late Triassic, 209-201 Ma | |
|---|---|
.jpg) | |
| Mounted skeleton cast at the Cleveland Museum of Natural History | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Order: | Saurischia |
| Suborder: | Theropoda |
| Family: | †Coelophysidae |
| Genus: | †Coelophysis |
| Species: | †C. bauri |
| Binomial name | |
| Coelophysis bauri (Cope, 1887) | |
| Synonyms | |
|
Coelurus bauri | |
Coelophysis bauri is an extinct species of coelophysid dinosaur that lived 209-201 million years ago[2] during the latter part of the Triassic Period in what is now the southwestern United States.[3] It is the type species of the genus Coelophysis, a group of small, slenderly-built, ground-dwelling, bipedal carnivores. It was originally classified in the genus Coelurus by Edward Drinker Cope in 1887.
Description
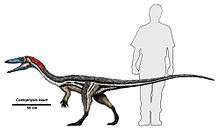
Coelophysis bauri is known from a number of complete fossil skeletons. C. bauri was a lightly built dinosaur which measured up to 3 metres (9.8 ft) in length[4] and which was more than a meter tall at the hips. Paul (1988) estimated the weight of the gracile form at 15 kg (33 lb), and the weight of the robust form at 20 kg (44 lb).[5] C. bauri was a bipedal, carnivorous, theropod dinosaur that was a fast and agile runner.[6] C. bauri had a furcula (wishbone), the earliest known example in a dinosaur.[7]
Coelophysis bauri had narrow hips, forelimbs adapted for grasping, and narrow feet.[8] Its neck and tail were long and slender.[3] The pelvis and hindlimbs of C. bauri are also slight variations on the theropod body plan. It has the open acetabulum and straight ankle hinge that define the Dinosauria. The hindlimb ended in a three-toed foot (pes), with a raised hallux. The tail had an unusual structure within its interlocking prezygapophysis of its vertebrae, which formed a semi-rigid lattice, apparently to stop the tail from moving up and down.[9]
Coelophysis bauri had a long narrow head (approximately 270 mm (0.9 ft)), with large, forward-facing eyes that afforded it stereoscopic vision and as a result excellent depth perception. Rinehart et al. (2004) described the complete sclerotic ring found for a juvenile C. bauri (specimen NMMNH P-4200), and compared it to data on the sclerotic rings of reptiles and birds and concluded that this Coelophysis was a diurnal, visually oriented predator."[10] The study found that its vision was superior to most lizards' vision, and ranked with that of modern birds of prey. The eyes appear to be the closest to those of eagles and hawks, with a high power of accommodation. The data also suggested poor night vision which would mean this dinosaur had a round rather than a split pupil.[6]
Coelophysis bauri had an elongated snout with large fenestrae which helped to reduce skull weight, while narrow struts of bones preserved the structural integrity of the skull. The neck had a pronounced sigmoid curve. The braincase is known, but little data could be derived because the skull was crushed.[8] Unlike some other theropods, the cranial ornamentation of Coelophysis bauri was not located at the top of its skull. Low, laterally raised bony ridges were present on the dorsolateral margin of the nasal and lacrimal bones in the skull, directly above the antorbital fenestra.[11]
History of discovery

Named in 1887, the type species of Coelophysis was originally named as a species of Coelurus.[12] Edward Drinker Cope first named Coelophysis in 1889 to name a new genus, outside of Coelurus and Tanystropheus to which C. bauri was previously classified in for C. bauri, C. willistoni and C. longicollis.[13] An amateur fossil collector working for Cope, David Baldwin, had found the first remains of the dinosaur in 1881 in the Chinle Formation in northwestern New Mexico.[14] Early in 1887 Cope referred the specimens collected to two new species, C. bauri and C. longicollis and erected the genus Coelurus. Later in 1887 Cope reassigned the material to a yet another genus, Tanystropheus. Two years later, Cope corrected his classification after realizing differences in the vertebrae, and named Coelophysis, with C. bauri as the type species,[13] which was named for Baur, another fossil collector who supplied Cope.[14] However, these first finds were too poorly preserved to give a complete picture of this new dinosaur. In 1947, a substantial 'graveyard' of Coelophysis fossils was found by George Whitaker, the assistant of Edwin H. Colbert, in New Mexico, at the Ghost Ranch, close to the original find. American Museum of Natural History paleontologist Edwin H. Colbert conducted a comprehensive study[11] of all the fossils found up to that date and assigned them to Coelophysis. The Ghost Ranch specimens were so numerous, including many well-preserved and fully articulated specimens, that one of them has since become the diagnostic, or type specimen, for the entire genus, replacing the original, poorly preserved specimen.[15] Coelophysis bauri was also found in the Siltstone Member of the Chinle Formation.
Classification

|
C. bauri is the type species of the genus Coelophysis and family Coelophysidae. Two contemporary species, C. longicollis and C. willistoni, are considered synonymous with C. bauri.[17]
In the early 1990s, there was debate over the diagnostic characteristics of the first specimens collected, compared to the material excavated at the Ghost Ranch Coelophysis quarry. Some paleontologists were of the opinion that the original specimens were not diagnostic beyond themselves and, therefore, that the name C. bauri could not be applied to any additional specimens. They therefore applied a different name, Rioarribasaurus colberti, to the Ghost Ranch quarry specimens.[18]
Since the numerous well-preserved Ghost Ranch specimes were used as Coelophysis bauri in most of the scientific literature, the use of Rioarribasaurus colberti would have been very inconvenient for researchers, so a petition was given to have the type specimen of Coelophysis transferred from the poorly preserved original specimen to one of the well-preserved Ghost ranch specimens. This would make Rioarribasaurus colberti a definite synonym of Coelophysis bauri, specifically a junior objective synonym.[12] In the end, the International Commission on Zoological Nomenclature (ICZN) voted to make one of the Ghost Ranch samples the actual type specimen for Coelophysis bauri and dispose of the name Rioarribasaurus colberti altogether (declaring it a nomen rejectum, or "rejected name"), thus resolving the confusion. The name Coelophysis bauri therefore became a nomen conservandum ("conserved name").[15]
Sullivan & Lucas (1999) renamed the type specimen from Cope's original material of Coelophysis bauri (specimen AMNH 2706) as Eucoelophysis baldwini.[19] Subsequent studies have shown that this original specimen was not a theropod at all, and is actually a silesaurid.[20]
Paleobiology
Feeding

The teeth of Coelophysis were typical of predatory dinosaurs, blade-like, recurved, sharp and jagged with fine serrations on both the anterior and posterior edges. Its dentition shows that it was carnivorous, probably preying on the small, lizard-like animals that were discovered with it.[21] It may also have hunted in packs to tackle larger prey.[5] Coelophysis bauri has approximately 26 teeth on maxillary bone of the upper jaw and 27 teeth on the dentary bone of the lower jaw.[11] Carpenter (2002) examined the bio-mechanics of theropods forelimbs and attempted to evaluate their usefulness in predation. He concluded that the forelimb of Coelophysis was flexible and had a good range of motion, but its bone structure suggested that it was comparatively weak. The "weak" forelimbs and small teeth in this genus, suggested that Coelophysis preyed upon animals that were substantially smaller than itself. Rinehart et al. agreed that Coelophysis was a "hunter of small, fast-moving prey".[6] Carpenter also identified three distinct models of theropod forelimb use and noted that Coelophysis was a "combination grasper-clutcher" as compared to other dinosaurs that were "clutchers" or "long armed graspers".[22]

It has been suggested that C. bauri was a cannibal, based on supposed juvenile specimens found "within" the abdominal cavities of some Ghost Ranch specimens.[23] However, Rob Gay showed in 2002 that these specimens were misinterpreted. Several specimens of "juvenile coelophysids" were actually small crurotarsan reptiles such as Hesperosuchus.[24] Gay's position was lent support in a 2006 study by Nesbitt et al.[25] In 2009, new evidence of cannibalism came to light when additional preparation of previously excavated matrix revealed regurgitate material in and around the mouth of Coelophysis specimen NMMNH P-44551. This material included tooth and jaw bone fragments that Rinehart et al. considered "morphologically identical" to a juvenile Coelophysis.[6]
In 2010, Gay examined the bones of juveniles found within the thoracic cavity of AMNH 7224, and calculated that the total volume of these bones was 17 times greater than the maximum estimated stomach volume of the Coelophysis specimen. Gay observed that the total volume would be even greater when considering that there would have been flesh on these bones. This analysis also noted the absence of tooth marks on the bones as would be expected in defleshing, and the absence of expected pitting by stomach acids. Finally Gay demonstrated that the alleged cannibalized juvenile bones, were deposited stratigraphically below the larger animal that supposedly cannibalized them. Taken together these data suggested that the Coelophysis specimen AMNH 7224 was not a cannibal and that the bones of the juvenile and adult specimens were found in their final position as a result of "coincidental superposition of different sized individuals.[26]
Pack behavior

The discovery of over 1000 specimens of Coelophysis at the Whitaker quarry at Ghost Ranch, has suggested gregarious behavior to researchers like Schwartz and Gillette.[27] There is a tendency to see this massive congregation of animals as evidence for huge packs of Coelophysis bauri roaming the land.[23] No direct evidence for flocking exists; the deposits only indicate that large numbers of Coelophysis, along with other Triassic animals, were buried together. Some of the evidence from the taphonomy of the site indicates that these animals may have been gathered together to feed or drink from a depleted water hole or to feed on a spawning run of fish, and then became buried in a catastrophic flash flood[23][27] or a drought.[23]
Growth and sexual dimorphism
Rinehart (2009) assessed the ontogenic growth of this genus using data gathered from the length of its upper leg bone (femur) and concluded that Coelophysis juveniles grew rapidly like birds (and not slowly like reptiles), especially during the first year of life.[6] Coelophysis likely reached sexual maturity between the second and third year of life and reached its full size, just above 10 feet in length, by its eighth year. This study identified four distinct growth stages: 1-year, 2-year, 4-year, and 7+ year.[6] It was also thought, that as soon as they were hatched, they would have to fend for themselves.[23]
Two "morphs" of Coelophysis have been identified: a more gracile form, as in specimen AMNH 7223, and a slightly more robust form, as in specimens AMNH 7224 and NMMNH P-42200. Skeletal proportions were different between these two forms;[28] the gracile form has a longer skull, a longer neck, shorter forelimbs, and has sacral neural spines that are fused; and the robust form has a shorter skull, a shorter neck, longer forelimbs, and unfused sacral neural spines.[11] Historically, many arguments have been made that this represents some sort of dimorphism in the population of Coelophysis, probably sexual dimorphism.[5][11][29][30] Raath agreed that dimorphism in Coelophysis is evidenced by the size and structure of the forelimb.[29] Rinehart et al. studied 15 individuals, and agreed that two morphs were present, even in juvenile specimens, and suggested that sexual dimorphism was present early in life, prior to sexual maturity. Rinehart concluded that the gracile form was female and the robust form was male based on differences in the sacral vertebrae of the gracile form, which allowed for greater flexibility for egg laying.[6] Further support for this position was provided by an analysis showing that each morph comprised 50% of the population, as would be expected in a 50/50 sex ratio.[31]
However, more recent research has found that C. bauri had highly variable growth between individuals, with some specimens being larger in their immature phase than smaller adults were when completely mature; this indicates that the supposed presence of distinct morphs is simply the result of individual variation. This highly variable growth was likely ancestral to dinosaurs but later lost, and may have given such early dinosaurs an evolutionary advantage in surviving harsh environmental challenges.[32]
Reproduction
Through the compilation and analysis of a database of nearly three dozen birds and reptiles, and comparison with existing data about the anatomy of Coelophysis Rinehart et al. (2009) drew the following conclusions. It was estimated that average egg of Coelophysis was 31-33.5 millimeters across its minor diameter, and that each female would lay between 24-26 eggs in each clutch. The evidence suggested that some parental care was necessary to nurture the relatively small hatchlings during the first year of life, where they would reach 1.5 meters in length by the end of their first growth stage. Coelophysis bauri invested as much energy in reproduction as other extinct reptiles of its approximate size.[6][33]
Paleopathology
In a 2001 study conducted by Bruce Rothschild and other paleontologists, 14 foot bones referred to Coelophysis were examined for signs of stress fracture, but none were found.[34]
Ecology

Most specimens of Coelophysis bauri have been recovered at the Whitaker quarry ("Coelophysis quarry") at Ghost Ranch, in the Chinle Formation of Arizona and New Mexico. The Whitaker quarry has been dated to the Late Norian-Rhaetian stages of the Late Triassic.[35][2][36][37]
Ghost Ranch was located close to the equator 200 million years ago, and had a warm, monsoon-like climate with heavy seasonal precipitation.[38]
The multitude of specimens deposited so closely together at Ghost Ranch was probably the result of a flash flood, which swept away a large number of Coelophysis and buried them quickly and simultaneously. In fact, it seems that such flooding was commonplace during this period of the Earth's history and, indeed, the Petrified Forest of nearby Arizona is the result of a preserved log jam of tree trunks that were caught in one such flood. Whitaker quarry at Ghost Ranch is considered a monotaxic site because it features multiple individuals of a single taxon. The quality of preservation and the ontogenic (age) range of the specimens helped make Coelophysis one of the best known of all genera.[39] In 2009, Rinehart et al. noted that in one case the Coelophysis specimens were "washed into a topographic low containing a small pond, where they probably drowned and were buried by a sheet flood event from a nearby river."[6]
References
- 1 2 3 4 Carrano, M.T.; Benson, R.B.J.; & Sampson, S.D. (2012). "The phylogeny of Tetanurae (Dinosauria: Theropoda)". Journal of Systematic Palaeontology. 10: 211–300. doi:10.1080/14772019.2011.630927.
- 1 2 Fong, RK, LeBlanc, AR, Berman, DS, Reisz, RR. (2016). "Dental histology of Coelophysis bauri and the evolution of tooth attachment tissues in early dinosaurs". Journal of Morphology. 277 (7): 916–924. PMID 27087142. doi:10.1002/jmor.20545.
- 1 2 Gaines, Richard M. (2001). Coelophysis. ABDO Publishing Company. p. 4. ISBN 1-57765-488-9.
- ↑ Schwartz, Hilde L.; Gillette, David D. (1994). "Geology and taphonomy of the Coelophysis quarry, Upper Triassic Chinle Formation, Ghost Ranch, New Mexico". Journal of Paleontology. 68 (5): 1118–1130. JSTOR 1306181.
- 1 2 3 Paul, Gregory S. (1988). Predatory Dinosaurs of the World. Simon & Schuster. p. 260. ISBN 0-671-61946-2.
- 1 2 3 4 5 6 7 8 9 Rinehart, L.F.; Lucas, S.G.; Heckert, A.B.; Spielmann, J.A. & Celesky, M.D. (2009). "The paleobiology of Coelophysis bauri (Cope) from the Upper Triassic (Apachean) Whitaker quarry, New Mexico, with detailed analysis of a single quarry block". New Mexico Museum of Natural History & Science, a division of the Department of Cultural Affairs Bulletin. 45: 260.
- ↑ Rinehart, L.F.; Lucas, S.G. & Hunt, A.P. (2007). "Furculae in the Late Triassic theropod dinosaur Coelophysis bauri". Paläontologische Zeitschrift. 81 (2): 174–180. doi:10.1007/bf02988391.
- 1 2 Tykoski, R.S. & Rowe, T. (2004). "Ceratosauria": In: Weishampel, D.B.; Dodson, P.; Osmolska, H. (eds.). The Dinosauria (Second ed.). Berkeley: University of California Press. pp. 47–70. ISBN 0-520-24209-2.
- ↑ Gay, R.J. (2001). "An unusual adaptation in the caudal vertebrae of Coelophysis bauri (Dinosauria: Theropoda)". PaleoBios. 21: 55.
- ↑ Rinehart, L.F.; Heckert, A.B.; Lucas, S.G. & Hunt, A.P. (2004). "The sclerotic ring of the Late Triassic theropod dinosaur Coelophysis". New Mexico Geological Society Spring Meeting. 26: 64.
- 1 2 3 4 5 Colbert, E. (1989). "The Triassic Dinosaur Coelophysis". Museum of Northern Arizona Bulletin. 57: 160.
- 1 2 Colbert, E.H.; Charig, A.J.; Dodson, P.; Gillette, D.D.; Ostrom, J.H. & Weishampel, D.B. (1992). "Coelurus bauri Cope, 1887 (currently Coelophysis bauri; Reptilia, Saurischia): Proposed replacement of the lectotype by a neotype" (PDF). Bulletin of Zoological Nomenclature. 49 (4): 276–279.
- 1 2 Cope, E.D. (1889). "On a new genus of Triassic Dinosauria". The American Naturalist. 23: 626. doi:10.1086/274979.
- 1 2 Glut, D.F. (1999). Dinosaurs, the Encyclopedia, Supplement 1. McFarland & Company, Inc. p. 442. ISBN 0-7864-0591-0.
- 1 2 International Commission on Zoological Nomenclature (1996). "Opinion 1842: Coelurus bauri Cope, 1887 (currently Coelophysis bauri; Reptilia, Saurischia): lectotype replaced by a neotype" (PDF). Bulletin of Zoological Nomenclature. 53 (2): 142–144.
- ↑ Ezcurra M. D., Brusatte S. L. (2011). "Taxonomic and phylogenetic reassessment of the early neotheropod dinosaur Camposaurus arizonensis from the Late Triassic of North America". Palaeontology. 54 (4): 763–772.
- ↑ Bristowe, A. & Raath, M.A. (2004). "A juvenile coelophysoid skull from the Early Jurassic of Zimbabwe, and the synonymy of Coelophysis and Syntarsus". Palaeontologia Africana. 40: 31–41.
- ↑ Hunt, A.P. & Lucas, S.G. (1991). "Rioarribasaurus, a new name for a Late Triassic dinosaur from New Mexico (USA)". Paläontologische Zeitschrift. 65 (1/2): 191–198. doi:10.1007/bf02985783.
- ↑ Sullivan, R.M. & Lucas, S.G. (1999). "Eucoelophysis baldwini, a new theropod dinosaur from the Upper Triassic of New Mexico, and the status of the original types of Coelophysis". Journal of Vertebrate Paleontology. 19 (1pages=81-90): 81–90. doi:10.1080/02724634.1999.10011124.
- ↑ Nesbitt, S.J.; Irmis, R.B. & Parker, W.G. (2007). "A critical re-evaluation of the Late Triassic dinosaur taxa of North America". Journal of Systematic Palaeontology. 5 (2): 209–243. doi:10.1017/S1477201907002040.
- ↑ Benton, M. (1993). Dinosaur and other prehistoric animal Fact Finder. Kingfisher Books Ldt. p. 256. ISBN 0-86272-949-1.
- ↑ Carpenter, K. (2002). "Forelimb bio-mechanics of non-avian theropod dinosaurs in predation". Senckenbergiana Lethaea. 82: 59–76. doi:10.1007/bf03043773.
- 1 2 3 4 5 Haines, T. & Chambers, P. (2007). The Complete Guide to Prehistoric Life. Firefly Books. pp. 70–71. ISBN 978-1-55407-181-4.
- ↑ Gay, R.J. (2002). "The myth of cannibalism in Coelophysis bauri". Journal of Vertebrate Paleontology. 22 (3): 57A.
- ↑ Nesbitt, S.J.; Turner, A.H.; Erickson, G.M. & Norell, M.A. (2006). "Prey choice and cannibalistic behaviour in the theropod Coelophysis". Biology Letters. 22. 2 (4): 611–614. PMC 1834007
 . PMID 17148302. doi:10.1098/rsbl.2006.0524.
. PMID 17148302. doi:10.1098/rsbl.2006.0524. - ↑ Gay, R.J. (2010a). Notes on Early Mesozoic Theropods (First ed.). Lulu press. pp. 9–24. ISBN 978-0-557-46616-0.
- 1 2 Schwartz, H.L. & Gillette, D.D. (1994). "Geology and taphonomy of the Coelophysis quarry, Upper Triassic Chinle Formation, Ghost Ranch, New Mexico". Journal of Paleontology. 68 (5): 1118–1130. JSTOR 1306181.
- ↑ Paul, Gregory S. (2010). The Princeton Field Guide to Dinosaurs. Princeton University Press. p. 74. ISBN 978-0-691-13720-9.
- 1 2 Carpenter, K. & Currie, P.J. (1990). Dinosaur Systematics: Approaches and Perspectives. Cambridge: Cambridge University Press. pp. 91–105. ISBN 0-521-43810-1.
- ↑ Gay, R. (2005). "Sexual Dimorphism in the Early Jurassic Theropod Dinosaur Dilophosaurus and a Comparison with Other Related Forms": In: Carpenter, K. (ed.). The Carnivorous Dinosaurs. Bloomington: Indiana University Press. pp. 277–283. ISBN 0-253-34539-1.
- ↑ Rinehart, L.F.; Lucas, S.G. & Heckert, A.B. (2001). "Preliminary statistical analysis defining the juvenile, robust and gracile forms of the Triassic dinosaur Coelophysis". Journal of Vertebrate Paleontology. 21: 93A. doi:10.1080/02724634.2001.10010852.
- ↑ Griffin, C.T.; Nesbitt, S.J. (2016). "Anomalously high variation in postnatal development is ancestral for dinosaurs but lost in birds". Proceedings of the National Academy of Sciences of the United States of America. 113: 14757–14762. doi:10.1073/pnas.1613813113.
- ↑ Glut, D.F. (2012). Dinosaurs, the Encyclopedia, Supplement 7. McFarland & Company, Inc. p. 866. ISBN 0-7864-4859-8.
- ↑ Rothschild, B.; Tanke, D.H. & Ford, T.L. (2001). "Theropod stress fractures and tendon avulsions as a clue to activity": In: Tanke, D. H.; Carpenter, K. (eds.). Mesozoic Vertebrate Life. Indiana University Press. pp. 331–336. ISBN 0-253-33907-3.
- ↑ Harris, Jereld D., Downs, Alex (2002). "A Drepanosaurid Pectoral Girdle from the Ghost Ranch (Whitaker) Coelophysis Quarry (Chinle Group, Rock Point Formation, Rhaetian), New Mexico". Journal of Vertabrate Paleontology. 22 (1): 70–75.
- ↑ Irmis R. B., Mundil R., Martz J. W., Parker W. G. (2011). "High-resolution U–Pb ages from the Upper Triassic Chinle Formation (New Mexico, USA) support a diachronous rise of dinosaurs". Earth and Planetary Science Letters. 309 (3): 258–267.
- ↑ Laing, Adam M., Turner, Alan H., Kernan, Ciara, Werning, Sarah, Smith, Nathan, Irmis, Randal, Nesbitt, Sterling J. (2017). "A New Shuvosaur Taxon (Archosauria, Pesudosuchia) from the Late Triassic Hayden Quarry of New Mexico, USA". Society of Vertebrate Paleontology 77th Annual Meeting Program & Abstracts:: 147.
- ↑ Braginetz, Donna (2007). "A new species of dinosauromorph (lower left) was among the mixed assemblage of dinosaurs and dinosauromorphs found at Hayden Quarry in Ghost Ranch, N.M.". American Museum of Natural History. Retrieved 31 March 2013.
- ↑ Glut, D.F. (2006). Dinosaurs, the Encyclopedia, Supplement 4. McFarland & Company, Inc. p. 749. ISBN 0-7864-2295-5.
| Wikimedia Commons has media related to Coelophysis. |
| Wikispecies has information related to: Coelophysis |
Avemetatarsalians (Dinosauromorpha and relatives) | |||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||
| |||||||||||||||||||||||||||||||




