Chlorophyll


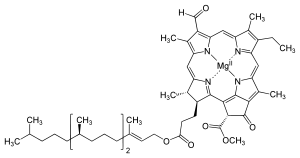
Chlorophyll (also chlorophyl) is any of several closely related green pigments found in cyanobacteria and the chloroplasts of algae and plants.[1] Its name is derived from the Greek words χλωρός, chloros ("green") and φύλλον, phyllon ("leaf").[2] Chlorophyll is essential in photosynthesis, allowing plants to absorb energy from light.
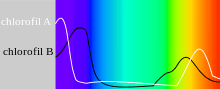
Chlorophyll absorbs light most strongly in the blue portion of the electromagnetic spectrum, followed by the red portion.[3] Conversely, it is a poor absorber of green and near-green portions of the spectrum, which it reflects, producing the green color of chlorophyll-containing tissues. Chlorophyll molecules are specifically arranged in and around photosystems that are embedded in the thylakoid membranes of chloroplasts. Two types of chlorophyll exist in the photosystems of green plants: chlorophyll a and b.[4]
Chlorophyll was first isolated and named by Joseph Bienaimé Caventou and Pierre Joseph Pelletier in 1817.[5]
Photosynthesis
Chlorophyll is vital for photosynthesis, which allows plants to absorb energy from light.[6]
Chlorophyll molecules are specifically arranged in and around photosystems that are embedded in the thylakoid membranes of chloroplasts.[7] In these complexes, chlorophyll serves two primary functions. The function of the vast majority of chlorophyll (up to several hundred molecules per photosystem) is to absorb light and transfer that light energy by resonance energy transfer to a specific chlorophyll pair in the reaction center of the photosystems. The two currently accepted photosystem units are photosystem II and photosystem I, which have their own distinct reaction centres, named P680 and P700, respectively. These centres are named after the wavelength (in nanometers) of their red-peak absorption maximum. The identity, function and spectral properties of the types of chlorophyll in each photosystem are distinct and determined by each other and the protein structure surrounding them. Once extracted from the protein into a solvent (such as acetone or methanol),[8][9][10] these chlorophyll pigments can be separated into chlorophyll a and chlorophyll b.
The function of the reaction center of chlorophyll is to absorb light energy and transfer it to other parts of the photosystem. The absorbed energy of the photon is transferred to an electron in a process called charge separation. The removal of the electron from the chlorophyll is an oxidation reaction. The chlorophyll donates the high energy electron to a series of molecular intermediates called an electron transport chain. The charged reaction center of chlorophyll (P680+) is then reduced back to its ground state by accepting an electron stripped from water. The electron that reduces P680+ ultimately comes from the oxidation of water into O2 and H+ through several intermediates. This reaction is how photosynthetic organisms such as plants produce O2 gas, and is the source for practically all the O2 in Earth's atmosphere. Photosystem I typically works in series with Photosystem II; thus the P700+ of Photosystem I is usually reduced as it accepts the electron, via many intermediates in the thylakoid membrane, by electrons coming, ultimately, from Photosystem II. Electron transfer reactions in the thylakoid membranes are complex, however, and the source of electrons used to reduce P700+ can vary.
The electron flow produced by the reaction center chlorophyll pigments is used to pump H+ ions across the thylakoid membrane, setting up a chemiosmotic potential used mainly in the production of ATP (stored chemical energy) or to reduce NADP+ to NADPH. NADPH is a universal agent used to reduce CO2 into sugars as well as other biosynthetic reactions.
Reaction center chlorophyll–protein complexes are capable of directly absorbing light and performing charge separation events without the assistance of other chlorophyll pigments, but the probability of that happening under a given light intensity is small. Thus, the other chlorophylls in the photosystem and antenna pigment proteins all cooperatively absorb and funnel light energy to the reaction center. Besides chlorophyll a, there are other pigments, called accessory pigments, which occur in these pigment–protein antenna complexes.
Chemical structure

Chlorophyll is a chlorin pigment, which is structurally similar to and produced through the same metabolic pathway as other porphyrin pigments such as heme. At the center of the chlorin ring is a magnesium ion. This was discovered in 1906,[11] and was the first time that magnesium had been detected in living tissue.[12] For the structures depicted in this article, some of the ligands attached to the Mg2+ center are omitted for clarity. The chlorin ring can have several different side chains, usually including a long phytol chain. There are a few different forms that occur naturally, but the most widely distributed form in terrestrial plants is chlorophyll a. After initial work done by German chemist Richard Willstätter spanning from 1905 to 1915, the general structure of chlorophyll a was elucidated by Hans Fischer in 1940. By 1960, when most of the stereochemistry of chlorophyll a was known, Robert Burns Woodward published a total synthesis of the molecule.[12][13] In 1967, the last remaining stereochemical elucidation was completed by Ian Fleming,[14] and in 1990 Woodward and co-authors published an updated synthesis.[15] Chlorophyll f was announced to be present in cyanobacteria and other oxygenic microorganisms that form stromatolites in 2010;[16][17] a molecular formula of C55H70O6N4Mg and a structure of (2-formyl)-chlorophyll a were deduced based on NMR, optical and mass spectra.[18] The different structures of chlorophyll are summarized below:
| Chlorophyll a | Chlorophyll b | Chlorophyll c1 | Chlorophyll c2 | Chlorophyll d | Chlorophyll f | |
|---|---|---|---|---|---|---|
| Molecular formula | C55H72O5N4Mg | C55H70O6N4Mg | C35H30O5N4Mg | C35H28O5N4Mg | C54H70O6N4Mg | C55H70O6N4Mg |
| C2 group | -CH3 | -CH3 | -CH3 | -CH3 | -CH3 | -CHO |
| C3 group | -CH=CH2 | -CH=CH2 | -CH=CH2 | -CH=CH2 | -CHO | -CH=CH2 |
| C7 group | -CH3 | -CHO | -CH3 | -CH3 | -CH3 | -CH3 |
| C8 group | -CH2CH3 | -CH2CH3 | -CH2CH3 | -CH=CH2 | -CH2CH3 | -CH2CH3 |
| C17 group | -CH2CH2COO-Phytyl | -CH2CH2COO-Phytyl | -CH=CHCOOH | -CH=CHCOOH | -CH2CH2COO-Phytyl | -CH2CH2COO-Phytyl |
| C17-C18 bond | Single (chlorin) |
Single (chlorin) |
Double (porphyrin) |
Double (porphyrin) |
Single (chlorin) |
Single (chlorin) |
| Occurrence | Universal | Mostly plants | Various algae | Various algae | Cyanobacteria | Cyanobacteria |
 Structure of chlorophyll a |
 Structure of chlorophyll b |
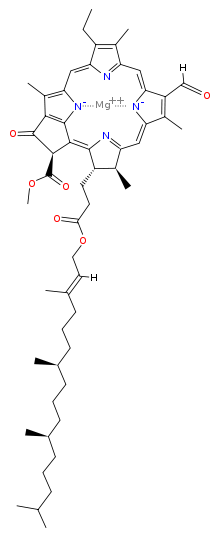 Structure of chlorophyll d |
 Structure of chlorophyll c1 |
 Structure of chlorophyll c2 |
 Structure of chlorophyll f |
When leaves degreen in the process of plant senescence, chlorophyll is converted to a group of colourless tetrapyrroles known as nonfluorescent chlorophyll catabolites (NCC's) with the general structure:
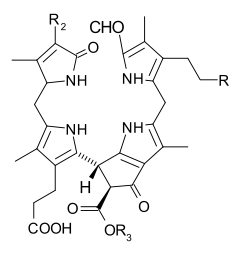
These compounds have also been identified in several ripening fruits.[19]
Measurement of chlorophyll content


Measurement of the absorption of light is complicated by the solvent used to extract the chlorophyll from plant material, which affects the values obtained,
- In diethyl ether, chlorophyll a has approximate absorbance maxima of 430 nm and 662 nm, while chlorophyll b has approximate maxima of 453 nm and 642 nm.[20]
- The absorption peaks of chlorophyll a are at 665 nm and 465 nm. Chlorophyll a fluoresces at 673 nm (maximum) and 726 nm. The peak molar absorption coefficient of chlorophyll a exceeds 105 M−1 cm−1, which is among the highest for small-molecule organic compounds.[21]
- In 90% acetone-water, the peak absorption wavelengths of chlorophyll a are 430 nm and 664 nm; peaks for chlorophyll b are 460 nm and 647 nm; peaks for chlorophyll c1 are 442 nm and 630 nm; peaks for chlorophyll c2 are 444 nm and 630 nm; peaks for chlorophyll d are 401 nm, 455 nm and 696 nm.[22]
By measuring the absorption of light in the red and far red regions, it is possible to estimate the concentration of chlorophyll within a leaf.[23]
Ratio fluorescence emission can be used to measure chlorophyll content. By exciting chlorophyll “a” fluorescence at a lower wavelength, the ratio of chlorophyll fluorescence emission at 705 nm +/- 10 nm and 735 nm +/-10 nm can provide a linear relationship of chlorophyll content when compared to chemical testing. The ratio F735/F700 provided a correlation value of r2 0.96 compared to chemical testing in the range from 41 mg m−2 up to 675 mg m−2. Gitelson also developed a formula for direct readout of chlorophyll content in mg m−2. The formula provided a reliable method of measuring chlorophyll content from 41 mg m−2 up to 675 mg m−2 with a correlation r2 value of 0.95.[24]
Biosynthesis
In plants, chlorophyll may be synthesized from succinyl-CoA and glycine, although the immediate precursor to chlorophyll a and b is protochlorophyllide. In Angiosperm plants, the last step, the conversion of protochlorophyllide to chlorophyll, is light-dependent and such plants are pale (etiolated) if grown in darkness. Non-vascular plants and green algae have an additional light-independent enzyme and grow green even in darkness.
Chlorophyll itself is bound to proteins and can transfer the absorbed energy in the required direction. Protochlorophyllide occurs mostly in the free form and, under light conditions, acts as a photosensitizer, forming highly toxic free radicals. Hence, plants need an efficient mechanism of regulating the amount of chlorophyll precursor. In angiosperms, this is done at the step of aminolevulinic acid (ALA), one of the intermediate compounds in the biosynthesis pathway. Plants that are fed by ALA accumulate high and toxic levels of protochlorophyllide; so do the mutants with the damaged regulatory system.[25]
Chlorosis is a condition in which leaves produce insufficient chlorophyll, turning them yellow. Chlorosis can be caused by a nutrient deficiency of iron—called iron chlorosis—or by a shortage of magnesium or nitrogen. Soil pH sometimes plays a role in nutrient-caused chlorosis; many plants are adapted to grow in soils with specific pH levels and their ability to absorb nutrients from the soil can be dependent on this.[26] Chlorosis can also be caused by pathogens including viruses, bacteria and fungal infections, or sap-sucking insects.
Complementary light absorbance of anthocyanins with chlorophylls
.PNG)
Anthocyanins are other plant pigments. The absorbance pattern responsible for the red color of anthocyanins may be complementary to that of green chlorophyll in photosynthetically active tissues such as young Quercus coccifera leaves. It may protect the leaves from attacks by plant eaters that may be attracted by green color.[27]
Distribution
The chlorophyll maps show milligrams of chlorophyll per cubic meter of seawater each month. Places where chlorophyll amounts were very low, indicating very low numbers of phytoplankton, are blue. Places where chlorophyll concentrations were high, meaning many phytoplankton were growing, are yellow. The observations come from the Moderate Resolution Imaging Spectroradiometer (MODIS) on NASA's Aqua satellite. Land is dark gray, and places where MODIS could not collect data because of sea ice, polar darkness, or clouds are light gray.The highest chlorophyll concentrations, where tiny surface-dwelling ocean plants are thriving, are in cold polar waters or in places where ocean currents bring cold water to the surface, such as around the equator and along the shores of continents. It is not the cold water itself that stimulates the phytoplankton. Instead, the cool temperatures are often a sign that the water has welled up to the surface from deeper in the ocean, carrying nutrients that have built up over time. In polar waters, nutrients accumulate in surface waters during the dark winter months when plants cannot grow. When sunlight returns in the spring and summer, the plants flourish in high concentrations.[28]
Culinary use
Chlorophyll is registered as a food additive (colorant), and its E number is E140. Chefs use chlorophyll to color a variety of foods and beverages green, such as pasta and absinthe.[29] Chlorophyll is not soluble in water, and it is first mixed with a small quantity of vegetable oil to obtain the desired solution.
See also
| Wikimedia Commons has media related to Chlorophyll. |
- Bacteriochlorophyll, related compounds in phototrophic bacteria
- Chlorophyll a, an essential chlorophyll pigment
- Chlorophyll b, also an essential chlorophyll pigment
- Chlorophyllin, a semi-synthetic derivative of chlorophyll
- Deep chlorophyll maximum
- Grow light, a lamp that promotes photosynthesis
- Chlorophyll fluorescence, to measure plant stress
References
- ↑ May, Paul. "Chlorophyll". University of Bristol.
- ↑ "chlorophyll". Online Etymology Dictionary.
- ↑ Muneer, Sowbiya; Kim, Eun Jeong; Park, Jeong Suk; Lee, Jeong Hyun (2014-03-17). "Influence of Green, Red and Blue Light Emitting Diodes on Multiprotein Complex Proteins and Photosynthetic Activity under Different Light Intensities in Lettuce Leaves (Lactuca sativa L.)". International Journal of Molecular Sciences. 15 (3): 4657–4670. ISSN 1422-0067. PMC 975419
 . PMID 24642884. doi:10.3390/ijms15034657.
. PMID 24642884. doi:10.3390/ijms15034657. - ↑ Speer, Brian R. (1997). "Photosynthetic Pigments". UCMP Glossary (online). University of California Museum of Paleontology. Retrieved 2010-07-17.
- ↑ See:
- Delépine, Marcel (September 1951). "Joseph Pelletier and Joseph Caventou". Journal of Chemical Education. 28 (9): 454. Bibcode:1951JChEd..28..454D. doi:10.1021/ed028p454.
- Pelletier and Caventou (1817) "Notice sur la matière verte des feuilles" (Notice on the green material in leaves), Journal de Pharmacie, 3 : 486-491. On p. 490, the authors propose a new name for chlorophyll. From p. 490: "Nous n'avons aucun droit pour nommer une substance connue depuis long-temps, et à l'histoire de laquelle nous n'avons ajouté que quelques faits ; cependant nous proposerons, sans y mettre aucune importance, le nom de chlorophyle, de chloros, couleur, et φυλλον, feuille : ce nom indiquerait le rôle qu'elle joue dans la nature." (We have no right to name a substance [that has been] known for a long time, and to whose story we have added only a few facts ; however, we will propose, without giving it any importance, the name chlorophyll, from chloros, color, and φυλλον, leaf : this name would indicate the role that it plays in nature.)
- ↑ Carter, J. Stein (1996). "Photosynthesis". University of Cincinnati. Archived from the original on 2013-06-29.
- ↑ Nature (July 5, 2013). "Unit 1.3. Photosynthetic Cells". Essentials of Cell Biology. nature.com.
- ↑ Marker, A. F. H. (1972). "The use of acetone and methanol in the estimation of chlorophyll in the presence of phaeophytin". Freshwater Biology. 2 (4): 361–385. doi:10.1111/j.1365-2427.1972.tb00377.x.
- ↑ Jeffrey, S. W.; Shibata, Kazuo (February 1969). "Some Spectral Characteristics of Chlorophyll c from Tridacna crocea Zooxanthellae". Biological Bulletin. Marine Biological Laboratory. 136 (1): 54–62. JSTOR 1539668. doi:10.2307/1539668.
- ↑ Gilpin, Linda (21 March 2001). "Methods for analysis of benthic photosynthetic pigment". School of Life Sciences, Napier University. Archived from the original on April 14, 2008. Retrieved 2010-07-17.
- ↑ Willstätter, Richard (1906) "Zur Kenntniss der Zusammensetzung des Chlorophylls" ([Contribution] to the knowledge of the composition of chlorophyll), Annalen der Chemie, 350 : 48-82. From p. 49: "Das Hauptproduct der alkalischen Hydrolyse bilden tiefgrüne Alkalisalze. In ihnen liegen complexe Magnesiumverbindungen vor, die das Metall in einer gegen Alkali auch bei hoher Temperatur merkwürdig widerstandsfähigen Bindung enthalten." (Deep green alkali salts form the main product of alkali hydrolysis. In them, complex magnesium compounds are present, which contain the metal in a bond that's extraordinarily resistant to alkali even at high temperature.)
- 1 2 Motilva, Maria-José (2008). "Chlorophylls – from functionality in food to health relevance". 5th Pigments in Food congress- for quality and health (Print). University of Helsinki. ISBN 978-952-10-4846-3.
- ↑ Woodward, R. B.; Ayer, W. A.; Beaton, J. M.; Bickelhaupt, F.; Bonnett, R.; Buchschacher, P.; Closs, G. L.; Dutler, H.; Hannah, J.; et al. (July 1960). "The total synthesis of chlorophyll". Journal of the American Chemical Society. 82 (14): 3800–3802. doi:10.1021/ja01499a093.
- ↑ Fleming, Ian (14 October 1967). "Absolute Configuration and the Structure of Chlorophyll". Nature. 216 (5111): 151–152. Bibcode:1967Natur.216..151F. doi:10.1038/216151a0.
- ↑ Woodward, R. B.; Ayer, William A.; Beaton, John M.; Bickelhaupt, Friedrich; Bonnett, Raymond; Buchschacher, Paul; Closs, Gerhard L.; Dutler, Hans; Hannah, John; et al. (1990). "The total synthesis of chlorophyll a" (PDF). Tetrahedron. 46 (22): 7599–7659. doi:10.1016/0040-4020(90)80003-Z.
- ↑ Jabr, Ferris (August 19, 2010) A New Form of Chlorophyll?. Scientific American. Retrieved on 2012-04-15.
- ↑ Infrared chlorophyll could boost solar cells. New Scientist. August 19, 2010. Retrieved on 2012-04-15.
- ↑ Chen, Min; Schliep, Martin; Willows, Robert D.; Cai, Zheng-Li; Neilan, Brett A.; Scheer, Hugo (September 2010). "A Red-Shifted Chlorophyll". Science. 329 (5997): 1318–1319. Bibcode:2010Sci...329.1318C. PMID 20724585. doi:10.1126/science.1191127.
- ↑ Müller, Thomas; Ulrich, Markus; Ongania, Karl-Hans; Kräutler, Bernhard (2007). "Colorless Tetrapyrrolic Chlorophyll Catabolites Found in Ripening Fruit Are Effective Antioxidants". Angewandte Chemie. 46 (45): 8699–8702. PMC 2912502 . PMID 17943948. doi:10.1002/anie.200703587.
- ↑ Gross, Jeana (1991). Pigments in vegetables: chlorophylls and carotenoids. Van Nostrand Reinhold, ISBN 0442006578.
- ↑ Porra, R. J. (1989). "Determination of accurate extinction coefficients and simultaneous equations for assaying chlorophylls a and b extracted with four different solvents: verification of the concentration of chlorophyll standards by atomic absorption spectroscopy". Biochimica et Biophysica Acta (BBA) - Bioenergetics. Volume 975 (3): 384, 394. doi:10.1016/S0005-2728(89)80347-0 – via Elsevier Science Direct.
- ↑ Larkum, edited by Anthony W. D. Larkum, Susan E. Douglas & John A. Raven (2003). Photosynthesis in algae. London: Kluwer. ISBN 0-7923-6333-7.
- ↑ Cate, Thomas; Perkins, T. D. (September 2003). "Joseph Pelletier and Joseph Caventou". Journal of Tree Physiology. 23 (15): 1077–1079. doi:10.1093/treephys/23.15.1077.
- ↑ Gitelson A. A., Buschmann C., Lichtenthaler H. K. (1999) “The Chlorophyll Fluorescence Ratio F735/F700 as an Accurate Measure of Chlorophyll Content in Plants” Remote Sens. Enviro. 69:296-302 (1999)
- ↑ Meskauskiene R; Nater M; Goslings D; Kessler F; op den Camp R; Apel K. (23 October 2001). "FLU: A negative regulator of chlorophyll biosynthesis in Arabidopsis thaliana". Proceedings of the National Academy of Sciences. 98 (22): 12826–12831. Bibcode:2001PNAS...9812826M. JSTOR 3056990. PMC 60138 . PMID 11606728. doi:10.1073/pnas.221252798.
- ↑ Duble, Richard L. "Iron Chlorosis in Turfgrass". Texas A&M University. Retrieved 2010-07-17.
- ↑ Karageorgou, P.; Manetas, Y. (2006). "The importance of being red when young: Anthocyanins and the protection of young leaves of Quercus coccifera from insect herbivory and excess light". Tree Physiology. 26 (5): 613–21. PMID 16452075. doi:10.1093/treephys/26.5.613.
- ↑ Chlorophyll : Global Maps. Earthobservatory.nasa.gov. Retrieved on 2014-02-02.
- ↑ Adams, Jad (2004). Hideous absinthe : a history of the devil in a bottle. United Kingdom: I.B.Tauris, 2004. p. 22. ISBN 1860649203.
| Subdisciplines | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Plant groups | |||||||||||
| |||||||||||
| |||||||||||
| Plant growth and habit | |||||||||||
| Reproduction | |||||||||||
| Plant taxonomy | |||||||||||
| Practice | |||||||||||
| |||||||||||
| |||||||||||