Catechol-''O''-methyl transferase

| catechol-O-methyltransferase | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Identifiers | |||||||||
| EC number | 2.1.1.6 | ||||||||
| CAS number | 9012-25-3 | ||||||||
| Databases | |||||||||
| IntEnz | IntEnz view | ||||||||
| BRENDA | BRENDA entry | ||||||||
| ExPASy | NiceZyme view | ||||||||
| KEGG | KEGG entry | ||||||||
| MetaCyc | metabolic pathway | ||||||||
| PRIAM | profile | ||||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||||
| Gene Ontology | AmiGO / EGO | ||||||||
| |||||||||

Catechol-O-methyltransferase (COMT; EC 2.1.1.6) is one of several enzymes that degrade catecholamines (such as dopamine, epinephrine, and norepinephrine), catecholestrogens, and various drugs and substances having a catechol structure.[5] In humans, catechol-O-methyltransferase protein is encoded by the COMT gene.[6] Two isoforms of COMT are produced: the soluble short form (S-COMT) and the membrane bound long form (MB-COMT). As the regulation of catecholamines is impaired in a number of medical conditions, several pharmaceutical drugs target COMT to alter its activity and therefore the availability of catecholamines.[7] COMT was first discovered by the biochemist Julius Axelrod in 1957.[8]
Function
Catechol-O-methyltransferase is involved in the inactivation of the catecholamine neurotransmitters (dopamine, epinephrine, and norepinephrine). The enzyme introduces a methyl group to the catecholamine, which is donated by S-adenosyl methionine (SAM). Any compound having a catechol structure, like catecholestrogens and catechol-containing flavonoids, are substrates of COMT.
Levodopa, a precursor of catecholamines, is an important substrate of COMT. COMT inhibitors, like entacapone, save levodopa from COMT and prolong the action of levodopa.[9] Entacapone is a widely used adjunct drug of levodopa therapy. When given with an inhibitor of dopa decarboxylase (carbidopa or benserazide), levodopa is optimally saved. This "triple therapy" is becoming a standard in the treatment of Parkinson's disease.
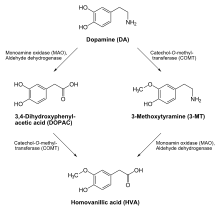
Specific reactions catalyzed by COMT include:
- Dopamine → 3-Methoxytyramine
- DOPAC → HVA (homovanillic acid)
- Norepinephrine → Normetanephrine
- Epinephrine → Metanephrine
- Dihydroxyphenylethylene glycol (DOPEG) → Methoxyhydroxyphenylglycol (MOPEG)
- 3,4-Dihydroxymandelic acid (DOMA) → Vanillylmandelic acid (VMA)
In the brain, COMT-dependent dopamine degradation is of particular importance in brain regions with low expression of the presynaptic dopamine transporter (DAT), such as the prefrontal cortex.[10][11][12][13] (In the PFC, dopamine is also removed by presynaptic norepinephrine transporters (NET) and degraded by monoamine oxidase.)[14] Controversy exists about the predominance and orientation of membrane bound COMT in the CNS,[15][16][17] that is, whether this COMT process is active intracellularly in postsynaptic neurons and glia, or oriented outward on the membrane, acting extracellularly on synaptic and extrasynaptic dopamine.
Soluble COMT can also be found extracellularly, although extracellular COMT plays a less significant role in the CNS than it does peripherally.[18]:210 Despite its importance in neurons, COMT is actually primarily expressed in the liver.[18]:135
Genetics in humans
The COMT protein is coded by the gene COMT. The gene is associated with allelic variants. The best-studied is Val158Met.[13] Others are rs737865 and rs165599 that have been studied, e.g., for association with personality traits,[19] response to antidepressant medications,[20] and psychosis risk associated with Alzheimer's disease.[21]
Val158Met polymorphism
A functional single-nucleotide polymorphism (a common normal variant) of the gene for catechol-O-methyltransferase results in a valine to methionine mutation at position 158 (Val158Met) rs4680.[13] In vitro, the homozygous Val variant metabolizes dopamine at up to four times the rate of its methionine counterpart.[20] However, in vivo the Met variant is overexpressed in the brain,[22] resulting in a 40% decrease (rather than 75% decrease) in functional enzyme activity.[23] The lower rates of catabolisis for the Met allele results in higher synaptic dopamine levels following neurotransmitter release, ultimately increasing dopaminergic stimulation of the post-synaptic neuron. Given the preferential role of COMT in prefrontal dopamine degradation, the Val158Met polymorphism is thought to exert its effects on cognition by modulating dopamine signaling in the frontal lobes.
The gene variant has been shown to affect cognitive tasks broadly related to executive function, such as set shifting, response inhibition, abstract thought, and the acquisition of rules or task structure.[24][25][26]
Comparable effects on similar cognitive tasks, the frontal lobes, and the neurotransmitter dopamine have also all been linked to schizophrenia. It has been proposed that an inherited variant of COMT is one of the genetic factors that may predispose someone to developing schizophrenia later in life, naturally or due to adolescent-onset cannabis use.[27] However, a more recent study cast doubt on the proposed connection between this gene and the effects of cannabis on schizophrenia development.[28]
It is increasingly recognised that allelic variation at the COMT gene are also relevant for emotional processing, as they seem to influence the interaction between prefrontal and limbic regions. Research conducted at the Section of Neurobiology of Psychosis, Institute of Psychiatry, King's College London has demonstrated an effect of COMT both in patients with bipolar disorder and in their relatives,[29] but these findings have not been replicated so far.
The COMT Val158Met polymorphism also has a pleiotropic effect on emotional processing.[29][30] Furthermore, the polymorphism has been shown to affect ratings of subjective well-being. When 621 women were measured with experience sample monitoring, which is similar to mood assessment as response to beeping watch, the met/met form confers double the subjective mental sensation of well-being from a wide variety of daily events. The ability to experience reward increased with the number of ‘Met’ alleles.[31] Also, the effect of different genotype was greater for events that were felt as more pleasant. The effect size of genotypic moderation was quite large: Subjects with the val/val genotype generated almost similar amounts of subjective well-being from a ‘very pleasant event’ as met/met subjects did from a ‘bit pleasant event’. Genetic variation with functional impact on cortical dopamine tone has a strong influence on reward experience in the flow of daily life.[31] In one study participants with the met/met phenotype described an increase of positive affect twice as high in amplitude as participants with the val/val phenotype following very pleasant or pleasant events.[31]
One review found that those with Val/Val tended to be more extroverted, more novelty seeking and less neurotic than those with the Met/Met allele[32]
Temporomandibular joint dysfunction
Temporomandibular joint dysfunction (TMD) does not appear to be a classic genetic disorder, however variations in the gene that codes for COMT have been suggested to be responsible for inheritance of a predisposition to develop TMD during life.[33]
Nomenclature
COMT is the name given to the gene that codes for this enzyme. The O in the name stands for oxygen, not for ortho.
COMT inhibitors
COMT inhibitors include tolcapone, entacapone and opicapone, which are used in the treatment of Parkinson's disease.[34] Risk of liver toxicity and related digestive disorders restricts the use of tolcapone.[35]
See also
Additional images
References
- ↑ "Human PubMed Reference:".
- ↑ "Mouse PubMed Reference:".
- ↑ Flower R, Rang HP, Dale MM, Ritter JM (2007). "Figure 11-4". Rang & Dale's pharmacology (6th ed.). Edinburgh: Churchill Livingstone. ISBN 0-443-06911-5.
- ↑ Rang HP, Dale MM, Ritter JM, Flower RJ, Henderson G (2011). "Figure 14.4". Rang & Dale's Pharmacology. Student consult (7th ed.). Elsevier Health Sciences. ISBN 978-0-7020-4504-2.
- ↑ "Test ID: COMT: Catechol-O-Methyltransferase Genotype". www.mayomedicallaboratories.com. Mayo Clinic: Mayo Medical Laboratories. Retrieved November 16, 2016.
- ↑ Grossman MH, Emanuel BS, Budarf ML (April 1992). "Chromosomal mapping of the human catechol-O-methyltransferase gene to 22q11.1----q11.2". Genomics. 12 (4): 822–5. PMID 1572656. doi:10.1016/0888-7543(92)90316-K.
- ↑ Tai CH, Wu RM (February 2002). "Catechol-O-methyltransferase and Parkinson's disease". Acta Medica Okayama. 56 (1): 1–6. PMID 11873938.
- ↑ Axelrod J (August 1957). "O-Methylation of Epinephrine and Other Catechols in vitro and in vivo". Science. 126 (3270): 400–1. PMID 13467217. doi:10.1126/science.126.3270.400.
- ↑ Ruottinen HM, Rinne UK (November 1998). "COMT inhibition in the treatment of Parkinson's disease". Journal of Neurology. 245 (11 Suppl 3): P25–34. PMID 9808337.
Goetz CG (May 1998). "Influence of COMT inhibition on levodopa pharmacology and therapy". Neurology. 50 (5 Suppl 5): S26–30. PMID 9591519. doi:10.1212/WNL.50.5_Suppl_5.S26. - ↑ Brodal P (2016). "Chapter 5: Neurotransmitters and their receptors". The Central Nervous System. Oxford University Press. p. 75. ISBN 978-0-19-022896-5.
- ↑ Scheggia D, Sannino S, Scattoni ML, Papaleo F (May 2012). "COMT as a drug target for cognitive functions and dysfunctions". CNS & Neurological Disorders Drug Targets. 11 (3): 209–21. PMID 22483296. doi:10.2174/187152712800672481.
- ↑ Diaz-Asper CM, Weinberger DR, Goldberg TE (January 2006). "Catechol-O-methyltransferase polymorphisms and some implications for cognitive therapeutics". NeuroRx. 3 (1): 97–105. PMC 3593358
 . PMID 16490416. doi:10.1016/j.nurx.2005.12.010.
. PMID 16490416. doi:10.1016/j.nurx.2005.12.010. - 1 2 3 Schacht, Joseph P. (October 2016). "COMT val158met moderation of dopaminergic drug effects on cognitive function: A critical review". Pharmacogenomics Journal. 16 (5): 430–438. PMC 5028240 . PMID 27241058. doi:10.1038/tpj.2016.43.
- ↑ Juarez B, Han MH (September 2016). "Diversity of Dopaminergic Neural Circuits in Response to Drug Exposure". Neuropsychopharmacology. 41 (10): 2424–46. PMC 4987841 . PMID 26934955. doi:10.1038/npp.2016.32.
- ↑ Nissinen E, ed. (2010). Basic Aspects of Catechol-O-Methyltransferase and the Clinical Applications of its Inhibitors. Academic Press. p. 34. ISBN 978-0-12-381327-5 – via Google books.
- ↑ Chen J, Song J, Yuan P, Tian Q, Ji Y, Ren-Patterson R, Liu G, Sei Y, Weinberger DR (October 2011). "Orientation and cellular distribution of membrane-bound catechol-O-methyltransferase in cortical neurons: implications for drug development". The Journal of Biological Chemistry. 286 (40): 34752–60. PMC 3186432 . PMID 21846718. doi:10.1074/jbc.M111.262790.
The cellular distribution of MB-COMT in cortical neurons remains unclear and the orientation of MB-COMT on the cellular membrane is controversial.
- ↑ Schott BH, Frischknecht R, Debska-Vielhaber G, John N, Behnisch G, Düzel E, Gundelfinger ED, Seidenbecher CI (2010). "Membrane-Bound Catechol-O-Methyl Transferase in Cortical Neurons and Glial Cells is Intracellularly Oriented". Frontiers in Psychiatry. 1: 142. PMC 3059651 . PMID 21423451. doi:10.3389/fpsyt.2010.00142.
It has been a matter of debate whether in neural cells of the CNS the enzymatic domain of MB-COMT is oriented toward the cytoplasmic or the extracellular compartment.
- 1 2 Golan DE, Tashjian AH. Principles of pharmacology (3rd ed.). Philadelphia: Wolters Kluwer Health. ISBN 1-60831-270-4. OCLC 705260923.
- ↑ Gold MS, Blum K, Oscar-Berman M, Braverman ER (January 2014). "Low dopamine function in attention deficit/hyperactivity disorder: should genotyping signify early diagnosis in children?". Postgraduate Medicine. 126 (1): 153–77. PMC 4074363 . PMID 24393762. doi:10.3810/pgm.2014.01.2735.
- 1 2 Porcelli S, Drago A, Fabbri C, Gibiino S, Calati R, Serretti A (March 2011). "Pharmacogenetics of antidepressant response". Journal of Psychiatry & Neuroscience. 36 (2): 87–113. PMC 3044192 . PMID 21172166. doi:10.1503/jpn.100059.
- ↑ DeMichele-Sweet MA, Sweet RA (2010). "Genetics of psychosis in Alzheimer's disease: a review". Journal of Alzheimer's Disease. 19 (3): 761–80. PMC 2942073 . PMID 20157235. doi:10.3233/JAD-2010-1274.
- ↑ Zhu G, Lipsky RH, Xu K, Ali S, Hyde T, Kleinman J, Akhtar LA, Mash DC, Goldman D (December 2004). "Differential expression of human COMT alleles in brain and lymphoblasts detected by RT-coupled 5' nuclease assay". Psychopharmacology. 177 (1–2): 178–84. PMID 15290009. doi:10.1007/s00213-004-1938-z.
- ↑ Chen J, Lipska BK, Halim N, Ma QD, Matsumoto M, Melhem S, Kolachana BS, Hyde TM, Herman MM, Apud J, Egan MF, Kleinman JE, Weinberger DR (November 2004). "Functional analysis of genetic variation in catechol-O-methyltransferase (COMT): effects on mRNA, protein, and enzyme activity in postmortem human brain". American Journal of Human Genetics. 75 (5): 807–21. PMC 1182110 . PMID 15457404. doi:10.1086/425589.
- ↑ Bruder GE, Keilp JG, Xu H, Shikhman M, Schori E, Gorman JM, Gilliam TC (December 2005). "Catechol-O-methyltransferase (COMT) genotypes and working memory: associations with differing cognitive operations". Biological Psychiatry. 58 (11): 901–7. PMID 16043133. doi:10.1016/j.biopsych.2005.05.010.
- ↑ Robinson S, Goddard L, Dritschel B, Wisley M, Howlin P (December 2009). "Executive functions in children with autism spectrum disorders". Brain and Cognition. 71 (3): 362–8. PMID 19628325. doi:10.1016/j.bandc.2009.06.007.
- ↑ Diamond A, Briand L, Fossella J, Gehlbach L (January 2004). "Genetic and neurochemical modulation of prefrontal cognitive functions in children". The American Journal of Psychiatry. 161 (1): 125–32. PMID 14702260. doi:10.1176/appi.ajp.161.1.125.
- ↑ Caspi A, Moffitt TE, Cannon M, McClay J, Murray R, Harrington H, Taylor A, Arseneault L, Williams B, Braithwaite A, Poulton R, Craig IW (May 2005). "Moderation of the effect of adolescent-onset cannabis use on adult psychosis by a functional polymorphism in the catechol-O-methyltransferase gene: longitudinal evidence of a gene X environment interaction". Biological Psychiatry. 57 (10): 1117–27. PMID 15866551. doi:10.1016/j.biopsych.2005.01.026.
- ↑ Zammit S, Spurlock G, Williams H, Norton N, Williams N, O'Donovan MC, Owen MJ (November 2007). "Genotype effects of CHRNA7, CNR1 and COMT in schizophrenia: interactions with tobacco and cannabis use". The British Journal of Psychiatry. 191 (5): 402–7. PMID 17978319. doi:10.1192/bjp.bp.107.036129. Lay summary – MedWireNews.
- 1 2 Lelli-Chiesa G, Kempton MJ, Jogia J, Tatarelli R, Girardi P, Powell J, Collier DA, Frangou S (April 2011). "The impact of the Val158Met catechol-O-methyltransferase genotype on neural correlates of sad facial affect processing in patients with bipolar disorder and their relatives". Psychological Medicine. 41 (4): 779–88. PMID 20667170. doi:10.1017/S0033291710001431.
- ↑ Kempton MJ, Haldane M, Jogia J, Christodoulou T, Powell J, Collier D, Williams SC, Frangou S (April 2009). "The effects of gender and COMT Val158Met polymorphism on fearful facial affect recognition: a fMRI study". The International Journal of Neuropsychopharmacology / Official Scientific Journal of the Collegium Internationale Neuropsychopharmacologicum. 12 (3): 371–81. PMID 18796186. doi:10.1017/S1461145708009395.
- 1 2 3 Wichers M, Aguilera M, Kenis G, Krabbendam L, Myin-Germeys I, Jacobs N, Peeters F, Derom C, Vlietinck R, Mengelers R, Delespaul P, van Os J (December 2008). "The catechol-O-methyl transferase Val158Met polymorphism and experience of reward in the flow of daily life". Neuropsychopharmacology. 33 (13): 3030–6. PMID 17687265. doi:10.1038/sj.npp.1301520.
- ↑ Montag C, Jurkiewicz M, Reuter M (May 2012). "The role of the catechol-O-methyltransferase (COMT) gene in personality and related psychopathological disorders". CNS & Neurological Disorders Drug Targets. 11 (3): 236–50. PMC 4345409 . PMID 22483293. doi:10.2174/187152712800672382.
- ↑ Cairns BE (May 2010). "Pathophysiology of TMD pain--basic mechanisms and their implications for pharmacotherapy". Journal of Oral Rehabilitation. 37 (6): 391–410. PMID 20337865. doi:10.1111/j.1365-2842.2010.02074.x.
- ↑ Bonifácio MJ, Palma PN, Almeida L, Soares-da-Silva P (2007). "Catechol-O-methyltransferase and its inhibitors in Parkinson's disease". CNS Drug Reviews. 13 (3): 352–79. PMID 17894650. doi:10.1111/j.1527-3458.2007.00020.x.
- ↑ Jatana N, Apoorva N, Malik S, Sharma A, Latha N (January 2013). "Inhibitors of catechol-O-methyltransferase in the treatment of neurological disorders" (PDF). Central Nervous System Agents in Medicinal Chemistry. 13 (3): 166–94. PMID 24450388. doi:10.2174/1871524913666140109113341.
Two of the nitrocatechols, entacapone ... and tolcapone ... have been demonstrated to reduce the dose of L-DOPA required and also cause improvement in clinical symptoms, although tolcapone emerged to be more efficacious due to its greater bioavailability and a longer half-life when compared to entacapone. However, tolcapone is clinically restricted owning to its increased hepatotoxicity and other related digestive disorders.
Further reading
- Trendelenburg U (1991). "The interaction of transport mechanisms and intracellular enzymes in metabolizing systems". Journal of Neural Transmission. Supplementum. 32: 3–18. PMID 2089098. doi:10.1007/978-3-7091-9113-2_1.
- Zhu BT (October 2002). "On the mechanism of homocysteine pathophysiology and pathogenesis: a unifying hypothesis". Histology and Histopathology. 17 (4): 1283–91. PMID 12371153.
- Oroszi G, Goldman D (December 2004). "Alcoholism: genes and mechanisms". Pharmacogenomics. 5 (8): 1037–48. PMID 15584875. doi:10.1517/14622416.5.8.1037.
- Fan JB, Zhang CS, Gu NF, Li XW, Sun WW, Wang HY, Feng GY, St Clair D, He L (January 2005). "Catechol-O-methyltransferase gene Val/Met functional polymorphism and risk of schizophrenia: a large-scale association study plus meta-analysis". Biological Psychiatry. 57 (2): 139–44. PMID 15652872. doi:10.1016/j.biopsych.2004.10.018.
- Tunbridge EM, Harrison PJ, Weinberger DR (July 2006). "Catechol-o-methyltransferase, cognition, and psychosis: Val158Met and beyond". Biological Psychiatry. 60 (2): 141–51. PMID 16476412. doi:10.1016/j.biopsych.2005.10.024.
- Craddock N, Owen MJ, O'Donovan MC (May 2006). "The catechol-O-methyl transferase (COMT) gene as a candidate for psychiatric phenotypes: evidence and lessons". Molecular Psychiatry. 11 (5): 446–58. PMID 16505837. doi:10.1038/sj.mp.4001808.
- Frank MJ, Moustafa AA, Haughey HM, Curran T, Hutchison KE (October 2007). "Genetic triple dissociation reveals multiple roles for dopamine in reinforcement learning". Proceedings of the National Academy of Sciences of the United States of America. 104 (41): 16311–6. PMC 2042203 . PMID 17913879. doi:10.1073/pnas.0706111104.
External links
| Wikimedia Commons has media related to Catechol O-methyltransferase, COMT. |
- Catechol O-Methyltransferase at the US National Library of Medicine Medical Subject Headings (MeSH)