Brachiosaurus
| Brachiosaurus Temporal range: Late Jurassic, 154–153 Ma | |
|---|---|
 | |
| Reconstructed skeleton cast at the Field Museum of Natural History | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Dinosauria |
| Order: | Saurischia |
| Suborder: | †Sauropodomorpha |
| Clade: | †Sauropoda |
| Clade: | †Titanosauriformes |
| Family: | †Brachiosauridae |
| Genus: | †Brachiosaurus Riggs, 1903 |
| Type species | |
| †Brachiosaurus altithorax Riggs, 1903 | |
Brachiosaurus /ˌbrækiəˈsɔːrəs/ is a genus of sauropod dinosaur from the Jurassic Morrison Formation of North America. It was first described by Elmer S. Riggs in 1903 from fossils found in the Grand River Canyon (now Colorado River) of western Colorado, in the United States. Riggs named the dinosaur Brachiosaurus altithorax, declaring it "the largest known dinosaur". Brachiosaurus had a disproportionately long neck, small skull, and large overall size, all of which are typical for sauropods. However, the proportions of Brachiosaurus are unlike most sauropods: the forelimbs were longer than the hindlimbs, which resulted in a steeply inclined trunk, and its tail was shorter in proportion to its neck than other sauropods of the Jurassic.
Brachiosaurus is the namesake genus of the family Brachiosauridae, which includes a handful of other similar sauropods. Much of what is known by laypeople about Brachiosaurus is in fact based on Giraffatitan brancai, a species of brachiosaurid dinosaur from the Tendaguru Formation of Tanzania that was originally described by German paleontologist Werner Janensch as a species of Brachiosaurus. Recent research shows that the differences between the type species of Brachiosaurus and the Tendaguru material are significant enough that the African material should be placed in a separate genus. Several other potential species of Brachiosaurus have been described from Africa and Europe, but none of them are thought to belong to Brachiosaurus at this time.
Brachiosaurus is one of the rarer sauropods of the Morrison Formation. The type specimen of B. altithorax is still the most complete specimen, and only a relative handful of other specimens are thought to belong to the genus. It is regarded as a high browser, probably cropping or nipping vegetation as high as possibly 9 metres (30 ft) off of the ground. Unlike other sauropods, and its depiction in the film Jurassic Park, it was unsuited for rearing on its hindlimbs. It has been used as an example of a dinosaur that was most likely ectothermic because of its large size and the corresponding need for forage, but more recent research finds it to have been warm-blooded.
Description
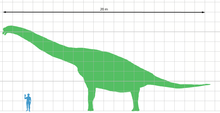
Because "Brachiosaurus" brancai (Giraffatitan) is known from much more complete material than B. altithorax, most size estimates for Brachiosaurus are actually for the African form. There is an additional element of uncertainty for North American Brachiosaurus because the most complete skeleton appears to have come from a subadult.[1] Over the years, the mass of B. altithorax has been estimated as 35.0 metric tons (38.6 short tons),[2] 43.9 metric tons (48.4 short tons),[3] 28.7 metric tons (31.6 short tons)[1] and, most recently, 56.3 metric tons (62.1 short tons).[4] In cases when the authors also provided estimates for Giraffatitan, and found that genus to be somewhat lighter (31.5 metric tons (34.7 short tons) for Paul [1988],[2] 39.5 metric tons (43.5 short tons) for Mazzetta et al [2004],[5] 23.3 metric tons (25.7 short tons) for Taylor [2009],[1] and 34.0 metric tons (37.5 short tons) for Benson et al [2014]).[4] The length of Brachiosaurus has been estimated at 26 metres (85 ft).[6]
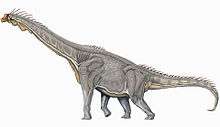
Like all sauropod dinosaurs, Brachiosaurus was a quadrupedal animal with a small skull, a long neck, a large trunk with a high-ellipsoid cross section, a long, muscular tail and slender, columnar limbs.[7] The skull had a robust, wide muzzle and thick jaw bones, with spoon–shaped teeth. As in Giraffatitan, there was an arch of bone over the snout and in front of the eyes that encircled the nasal opening, although this arch was not as large as in its relative.[8] Large air sacs connected to the lung system were present in the neck and trunk, invading the vertebrae and ribs, greatly reducing the overall density.[9][10] Unusually for a sauropod, the forelimbs were longer than the hind limbs. The humerus (upper arm bone) of Brachiosaurus was relatively lightly built for its size,[1] measuring 2.04 metres (6.7 ft) in length in the type specimen.[11] The femur (thigh bone) of the type specimen was only 2.03 metres (6.7 ft) long.[11] Unlike other sauropods, Brachiosaurus appears to have been slightly sprawled at the shoulder joint,[1] and the ribcage was unusually deep.[11] This led to the trunk being inclined, with the front much higher than the hips, and the neck exiting the trunk at a steep angle. Overall, this shape resembles a giraffe more than any other living animal.[2]
History of discovery

The genus Brachiosaurus, and type species B. altithorax, are based on a partial postcranial skeleton from Fruita, in the valley of the Colorado River of western Colorado.[12] This specimen was collected from rocks of the Brushy Basin Member of the Morrison Formation[13] in 1900 by Elmer S. Riggs and his crew from the Field Columbian Museum (now the Field Museum of Natural History) of Chicago.[11] It is currently cataloged as FMNH P 25107.[1] Riggs and company were working in the area as a result of favorable correspondence between Riggs and S. M. Bradbury, a dentist in nearby Grand Junction. In 1899, Riggs had sent inquiries to rural locations in the western United States concerning fossil finds, and Bradbury, an amateur collector himself, reported that dinosaur bones had been collected in the area since 1885.[12]

It was Riggs' field assistant H. W. Menke who found FMNH P 25107,[11] on July 4, 1900.[14] The locality, Riggs Quarry 13, was found on a small hill later known as Riggs Hill; it is marked by a plaque. Additional Brachiosaurus fossils are reported on Riggs Hill, but other fossil finds on the hill have been vandalized.[14][15] Riggs published a short report in 1901, noting the unusual length of the humerus compared to the femur and the extreme overall size and the resulting giraffe-like proportions, as well as the lesser development of the tail, but did not publish a name for the new dinosaur.[16] The titles of Riggs (1901) and (1903) suggested that the specimen was the largest known dinosaur.[11][16] Riggs followed his 1903 publication that named Brachiosaurus altithorax[11] with a more detailed description in a monograph in 1904.[17] Riggs derived the genus name from the Greek brachion/βραχιων meaning "arm" and sauros/σαυρος meaning "lizard", because he realized that the length of the arms was unusual for a sauropod.[11] The species epithet "altithorax" was chosen because of the unusually deep and wide chest cavity, from Latin altus meaning "deep" and Greek thorax/θώραξ (Latin thorax), meaning "breastplate, cuirass, corslet".[18]

The Fruita skeleton was not the first discovery of Brachiosaurus bones, although it was the first to be recognized as belonging to a new and distinct animal. In 1883, a sauropod skull was found near Garden Park, Colorado, at Felch Quarry 1, and was sent to Othniel Charles Marsh (of "Bone Wars" fame).[1] Marsh incorporated the skull into his skeletal restoration of Brontosaurus.[1][19] It eventually became part of the collections of the National Museum of Natural History, as USNM 5730.[1] In the 1970s, when Jack McIntosh and David Berman were working on the issue of the true skull of Apatosaurus, they reevaluated the Garden Park skull as more similar to Camarasaurus.[20] It was described and recognized as a Brachiosaurus skull in 1998 by Kenneth Carpenter and Virginia Tidwell, intermediate in form between Camarasaurus and Giraffatitan brancai (then still considered to be B. brancai).[21] Because there are no overlapping parts between this skull and FMNH P 25107, it cannot be confidently assigned to a species,[1][21] so it is classified as Brachiosaurus sp.[21]

Preparation of P 25107, the holotype of Brachiosaurus, began in the fall of 1900 shortly after it was collected by Elmer Riggs for the Field Museum of Natural History (Chicago). As the preparation of each bone was finished, it was put on display in a glass case in Hall 35 of the Fine Arts Palace of the Worlds Columbian Exposition, Field Museum's first home. All the bones were on display by 1908 when Field Museum's newly mounted Apatosaurus was unveiled. However, no mount was attempted because only 20% of the skeleton had been recovered. In 1993, the holotype bones were molded and cast, and the missing bones were sculpted based on Giraffatitan material in Berlin. This plastic skeleton was mounted and, in 1994, put on display at the north end of Stanley Field Hall, the main exhibit hall of the Field Museum's current building. The real bones of the holotype were put on exhibit in two large glass cases at either end of the mounted cast. The mount stood until 1999, when it was moved to the B Concourse of United Airlines' Terminal One in O'Hare International Airport to make room for the museum's newly acquired T. rex, "SUE".[22] At the same time, the Field Museum mounted a second plastic cast of the skeleton (designed for outside use) and it has been on display outside the museum on the NW terrace ever since. The only real bones currently on display are the humerus and two dorsals in the Mesozoic Hall of the Field Museum's Evolving Planet exhibit.[23]

Additional discoveries of Brachiosaurus material in North America have been uncommon and consist of a handful of bones. Material has been described from Colorado,[1][24][25][26] Oklahoma,[1][27] Utah,[1][24] and Wyoming,[1][3] and undescribed material has been mentioned from several other sites.[1][13] One of these specimens, a shoulder blade from Dry Mesa Quarry, Colorado, is one of the specimens at the center of the Supersaurus/Ultrasauros issue of the 1980s and 1990s. In 1985, James A. Jensen described disarticulated sauropod remains from the quarry as belonging to several taxa, including the new genera Supersaurus and Ultrasaurus,[28] the latter renamed Ultrasauros shortly thereafter because another sauropod already had the name.[29] Later study showed that the "ultrasaur" material mostly belonged to Supersaurus, although the shoulder blade did not. Because the holotype of Ultrasauros, a back vertebra, was one of the specimens that was actually from Supersaurus, the name Ultrasauros is a synonym of Supersaurus. The shoulder blade is now assigned to Brachiosaurus, but the species is uncertain.[1][25] In addition, the Dry Mesa "ultrasaur" was not as large as had been thought; the dimensions of the shoulder's coracoid bone indicate that the animal was smaller than Riggs' original specimen of Brachiosaurus.[1]
Species
Brachiosaurus altithorax

FMNH P 25107, the holotype of both the genus Brachiosaurus and the species B. altithorax, consists of the right humerus (upper arm bone), the right femur (thigh bone), the right ilium (a hip bone), the right coracoid (a shoulder bone), the sacrum (fused vertebrae of the hip), the last seven thoracic (trunk) and two caudal (tail) vertebrae, and a number of ribs.[1][11][16] Riggs described the coracoid as from the left side of the body,[11][17][16] but restudy has shown it to be a right coracoid.[1]
In 1969, during a study by Kingham, Brachiosaurus altithorax, along with species now assigned to other genera, were moved from the genus. Kingham found "B." atalaiensis, "B." brancai, and B. altithorax were referable to Astrodon creating many new species of Astrodon apart from the type. Kingham's views of brachiosaurid taxonomy have not been accepted by many authors.[30]
Other assigned species
- "B." atalaiensis: Originally described by de Lapparent and Zbyszewski,[31] this material's reference to Brachiosaurus was doubted by Upchurch, Barret and Dodson,[7] who listed it as an unnamed brachiosaurid, and placed in its own genus Lusotitan by Antunes and Mateus.[32] De Lapparent and Zbyszewski described a series of remains but did not designate a type specimen. Antunes and Mateus selected a partial postcranial skeleton (MIGM 4978, 4798, 4801–4810, 4938, 4944, 4950, 4952, 4958, 4964–4966, 4981–4982, 4985, 8807, 8793–87934) as a lectotype; this specimen includes 28 vertebrae, chevrons, ribs, a possible shoulder blade, humeri, forearm bones, partial left pelvis, lower leg bones, and part of the right ankle. The low neural spines, the prominent deltopectoral crest of the humerus (a muscle attachment site on the upper arm bone), the elongated humerus (very long and slender), and the long axis of the ilium tilted upward indicate that Lusotitan is a brachiosaurid.[32]
- "B." brancai: Janensch based his description on "Skelett S" (skeleton S) from Tendaguru,[33] but later realized that it comprised two partial individuals: S I and S II.[34] He at first did not designate them as a syntype series, but in 1935 made S I (MB.R.2180) the lectotype. Taylor in 2009, unaware of this action, proposed the larger and more complete S II (MB.R.2181) as the lectotype.[1] It includes, among other bones, several dorsal (trunk) vertebrae, the left scapula, both coracoids, both sternals (breastbones), both humeri, both ulna and radii (lower arm bones), a right hand, a partial left hand, both pubes (a hip bone) and the right femur, tibia and fibula (shank bones). Later Taylor realised that Janensch had in 1935 designated the smaller skeleton S I as the lectotype.[35][36] A re-assessment of the relation between the African and American brachiosaur material indicates that a separate generic name is warranted for the Tendaguru material, so "B." brancai has been moved to its own genus: Giraffatitan.[1][2]
- "B." fraasi: erected by Janensch in 1914, but later synonymized with "B." brancai;[34] this material now belongs to Giraffatitan.[1]
- "B." nougaredi: This species is known from fragmentary remains discovered in eastern Algeria, in the Sahara Desert. The present type material consists of a sacrum and some of the left metacarpals and phalanges. Found at the discovery site but not collected were partial bones of the left forearm, wrist bones, a right shin bone, and fragments that may have come from metatarsals.[37] de Lapparent, who described and named the material in 1960, reported the discovery locality as being in the Late Jurassic–age Taouratine Series (he assigned the rocks this age in part because of the presumed presence of Brachiosaurus).[37] This material was found disjointed over an area of several hundred meters,[37] and probably does not represent a single species.[38]
Referred material

Taylor (2009) lists a number of specimens referred to Brachiosaurus. These include some material, e.g., a humerus from Potter Creek and some Dry Mesa material (the latter partly described as Ultrasauros by Jensen), that are either clearly not brachiosaurid in origin, or at least not clearly referable to Brachiosaurus.[1] In contrast, a cervical (neck) vertebra and the skull mentioned above may belong to either B. altithorax or an as-yet unknown brachiosaurid from North America.[1] The cervical was found near Jensen, Utah, by Jensen,[24] and – if it belongs to Brachiosaurus – is one of a handful of neck vertebrae known for American brachiosaurids.[1] There is no unambiguous material of the skull, neck, anterior dorsal (forward trunk) region, distal (lower) limbs or feet.[1] More recently, Carballido et al. (2012) reported on a nearly complete postcranial skeleton of a juvenile sauropod (approximately 2 metres (6.6 ft) long) from the Morrison Formation of the Bighorn Basin, north-central Wyoming. This specimen was originally thought to belong to a diplodocid, but the authors reinterpreted it as representing a brachiosaurid, probably Brachiosaurus altithorax.[39]
There was ample material referred to "B." brancai in the collections of the Museum für Naturkunde Berlin, some of which was destroyed during World War II. Other material was transferred to other institution throughout Germany, some of which was also destroyed. Additional specimens are likely among the material collected by the British Museum of Natural History's Tendaguru expedition.[40] Much or all of this material probably belongs to Giraffatitan, although some may represent a new brachiosaurid.[41]
Separation of Giraffatitan

When describing the brachiosaurid material from Tendaguru in 1914, Janensch listed a number of differences and commonalities between them and B. altithorax.[33] In three further publications in 1929,[34] 1950 [42] and 1961[43] Janensch compared the two species in more detail, listing 13 putative shared characters.[1] Of these, however, only four appear to be valid, while six pertain to more inclusive groups than Brachiosauridae, and the rest are either difficult to assess or refer to material that is not Brachiosaurus.[1]
In 1988, Gregory Paul published a new reconstruction of the skeleton of "B." brancai, highlighting a number of differences in proportion between it and B. altithorax. Chief among them is a difference in the way the trunk vertebrae vary: they are fairly uniform in B. altithorax, but vary widely in the African material. Paul believed that the limb and girdle elements of both species were very similar, and therefore suggested to separate them not at genus, but only at subgenus level.[2]
Giraffatitan was raised to genus level by Olshevsky without comment.[29] A detailed study of all material, including the limb and girdle bones, by Michael Taylor in 2009 found that there are significant differences between Brachiosaurus altithorax and the Tendaguru material in all elements known from both species. Taylor found 26 distinct osteological (bone-based) characters, a larger difference than that between, e.g., Diplodocus and Barosaurus, and therefore argued that the African material should be placed in its own genus, Giraffatitan, as G. brancai.[1] An important difference between the two genera is the overall body shape, with Brachiosaurus having a 23% longer dorsal (trunk) vertebrate series and a 20 to 25% longer and also taller tail.[1]
Classification
Brachiosaurus is the namesake genus of Brachiosauridae.[17] Over the years, a number of sauropods have been assigned to Brachiosauridae, such as Astrodon, Bothriospondylus, Dinodocus, Pelorosaurus, Pleurocoelus, and Ultrasaurus,[44] but most of these are currently regarded as dubious or of uncertain placement.[7] A phylogenetic analysis of sauropods published in the description of Abydosaurus found that genus to form a clade with Brachiosaurus and Giraffatitan (included in Brachiosaurus).[45] A more recent analysis focused on possible Asian brachiosaurid material found a clade including Abydosaurus, Brachiosaurus, Cedarosaurus, Giraffatitan, and Paluxysaurus, but not Qiaowanlong, the putative Asian brachiosaurid.[46] Related genera include Lusotitan and Sauroposeidon.[7] Brachiosauridae is situated at the base of Titanosauriformes, a group of sauropods that also includes the titanosaurs.[46]

According to the revised diagnosis by Taylor, Brachiosaurus altithorax is diagnosed by a plethora of characters, many to be found on the dorsal (back) vertebrae.[1] Among the characters placing it in the family Brachiosauridae are a ratio of humerus length to femur length of at least 0.9 (i.e. the upper arm bone is at least nearly as long as the thigh bone), and a very flattened femur shaft (ratio ≥1.85).[1]
Cladogram of Brachiosauridae after D'Emic (2012).[30]
| Brachiosauridae |
| |||||||||||||||||||||||||||
Paleobiology
Neck position
In contrast to most other sauropods, brachiosaurids had an inclined back, due to their long forelimbs. Therefore, if the neck exited the body in a straight line, it already pointed upwards.[1][47][48][49] The exact angle is influenced by how the pectoral girdle is reconstructed, that is how the shoulder blades are placed on the ribcage.[47][48][49] The mobility of the neck was reconstructed as quite low by Stevens and Parrish,[47][48][49] while other researchers like Paul and Christian and Dzemski argued for more flexible necks.[2][50]
Feeding and diet
Brachiosaurus is thought to have been a high browser, feeding on foliage well above the ground. Even if it did not hold its neck near vertical, and instead had a straighter neck, its head height may still have been over 9 metres (30 ft) above the ground.[8][3] It probably fed mostly on foliage above 5 metres (16 ft). This does not preclude the possibility that it also fed lower at times, between 3 to 5 metres (9.8 to 16.4 ft) up.[8] Its diet likely consisted of ginkgos, conifers, tree ferns, and large cycads, with intake estimated at 200 to 400 kilograms (440 to 880 lb) of plant matter daily.[8] However, more recent studies estimate that ~240 kilograms (530 lb) of plant matter would have been sufficient to feed a 70 metric tons (77 short tons) sauropod,[51] so Brachiosaurus may have required only about 120 kilograms (260 lb) of fodder a day. Brachiosaur feeding involved simple up–and–down jaw motion. The teeth were arranged to shear material as they closed, and were probably used to crop and/or nip vegetation.[52]
It has repeatedly been suggested, e.g. in the movie Jurassic Park, that Brachiosaurus could rear into a bipedal or tripodal (with tail support) pose to feed.[2] However, a detailed physical modelling-based analysis of sauropod rearing capabilities by Heinrich Mallison showed that while many sauropods could rear, the unusual brachiosaurid body shape and limb length ratio made them exceptionally ill suited for rearing. The forward position of the center of mass would have led to problems with stability, and required unreasonably large forces in the hips to obtain an upright posture. Brachiosaurus would also have gained relatively little from rearing (only 33% more feeding height), compared to other sauropods, for which a bipedal pose may have tripled the feeding height.[53]
Metabolism
Like all sauropods, Brachiosaurus was homeothermic (maintaining a stable internal temperature) and endothermic (controlling body temperature through internal means), meaning that it was able to actively control its body temperature ("warm-blooded"), producing the necessary heat through a high basic metabolic rate of its cells.[54] In the past, Brachiosaurus has been used an example of a dinosaur for which endothermy is unlikely, because of the combination of great size (leading to overheating) and great caloric needs to fuel endothermy.[55] However, these calculations were based on incorrect assumptions about the available cooling surfaces (the large air sacs were not known), and a grossly inflated body mass. These inaccuracies resulted in the overestimation of heat production and the underestimation of heat loss.[54] The large nasal arch has been postulated as an adaptation for cooling the brain, as a surface for evaporative cooling of the blood.[55]
Paleoecology

With the removal of the East African Giraffatitan, Brachiosaurus is known only from the Morrison Formation of western North America.[1] The Morrison Formation is interpreted as a semiarid environment with distinct wet and dry seasons,[56][57] and flat floodplains.[56] Vegetation varied from gallery forests (river–lining forests in otherwise treeless settings) of conifers, tree ferns, and ferns, to fern savannas with rare Araucaria-like trees.[58] Several other sauropod genera were present in the Morrison Formation, with differing body proportions and feeding adaptations.[3] Among these were Apatosaurus, Barosaurus, Camarasaurus, Diplodocus, Haplocanthosaurus, and Supersaurus.[3][59] Brachiosaurus was one of the less abundant Morrison Formation sauropods. In a survey of over 200 fossil localities, John Foster reported 12 specimens of the genus, comparable to Barosaurus (13) and Haplocanthosaurus (12), but far fewer than Apatosaurus (112), Camarasaurus (179), and Diplodocus (98).[3] Brachiosaurus fossils are found only in the lower-middle part of the expansive Morrison Formation (stratigraphic zones 2-4), dated to about 154-153 million years ago,[60] unlike many other types of sauropod which have been found throughout the formation.[3]

The Morrison Formation records an environment and time dominated by gigantic sauropod dinosaurs.[61] Other dinosaurs known from the Morrison include the theropods Koparion, Stokesosaurus, Ornitholestes, Ceratosaurus, Allosaurus and Torvosaurus, the sauropods Apatosaurus, Camarasaurus, and Diplodocus, and the ornithischians Camptosaurus, Dryosaurus, Othnielia, Gargoyleosaurus and Stegosaurus.[62] Diplodocus is commonly found at the same sites as Apatosaurus, Allosaurus, Camarasaurus, and Stegosaurus.[63] Allosaurus, which accounting for 70 to 75% of theropod specimens and was at the top trophic level of the Morrison food web.[64] Many of the dinosaurs of the Morrison Formation are the same genera as those seen in Portuguese rocks of the Lourinha Formation (mainly Allosaurus, Ceratosaurus, Torvosaurus, and Stegosaurus), or have a close counterpart (Brachiosaurus and Lusotitan, Camptosaurus and Draconyx).[65] Other vertebrates that shared this paleoenvironment included ray-finned fishes, frogs, salamanders, turtles like Dorsetochelys, sphenodonts, lizards, terrestrial and aquatic crocodylomorphans such as Hoplosuchus, and several species of pterosaur like Harpactognathus and Mesadactylus. Shells of bivalves and aquatic snails are also common. The flora of the period has been revealed by fossils of green algae, fungi, mosses, horsetails, cycads, ginkgoes, and several families of conifers. Vegetation varied from river-lining forests of tree ferns, and ferns (gallery forests), to fern savannas with occasional trees such as the Araucaria-like conifer Brachyphyllum.[66]
Cultural significance
Brachiosaurus is one of the best-known dinosaurs amongst both paleontologists and the general public. A main belt asteroid, 1991 GX7, has been named 9954 Brachiosaurus in honor of the genus.[67][68] The genus has been featured in many films and television programs, most notably the Jurassic Park and Walking with Dinosaurs series. The digital model of Brachiosaurus used in Jurassic Park went on to become the starting point for the ronto models in the 1997 special edition of the science fiction film Star Wars Episode IV: A New Hope.[69]
References
- 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 Taylor, M.P. (2009). "A re-evaluation of Brachiosaurus altithorax Riggs 1903 (Dinosauria, Sauropoda) and its generic separation from Giraffatitan brancai (Janensh 1914)" (PDF). Journal of Vertebrate Paleontology. 29 (3): 787–806. doi:10.1671/039.029.0309.
- 1 2 3 4 5 6 7 Paul, G.S. (1988). "The brachiosaur giants of the Morrison and Tendaguru with a description of a new subgenus, Giraffatitan, and a comparison of the world's largest dinosaurs" (pdf). Hunteria. 2 (3).
- 1 2 3 4 5 6 7 Foster, J.R. (2003). Paleoecological analysis of the vertebrate fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain region, U.S.A. New Mexico Museum of Natural History and Science Bulletin, 23. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science.
- 1 2 Benson, R. B. J.; Campione, N. S. E.; Carrano, M. T.; Mannion, P. D.; Sullivan, C.; Upchurch, P.; Evans, D. C. (2014). "Rates of Dinosaur Body Mass Evolution Indicate 170 Million Years of Sustained Ecological Innovation on the Avian Stem Lineage". PLoS Biology. 12 (5): e1001853. PMC 4011683
 . PMID 24802911. doi:10.1371/journal.pbio.1001853.
. PMID 24802911. doi:10.1371/journal.pbio.1001853. - ↑ Mazzetta, G.V.; Christiansen, P.; Farina, R.A. (2004). "Giants and Bizarres: Body Size of Some Southern South American Cretaceous Dinosaurs". Historical Biology. 16 (2–4): 1–13. doi:10.1080/08912960410001715132.
- ↑ Holtz, Thomas R. Jr. (2008) Dinosaurs: The Most Complete, Up-to-Date Encyclopedia for Dinosaur Lovers of All Ages Supplementary Information
- 1 2 3 4 Upchurch, P., Barrett, P.M. & Dodson, P. (2004): "Sauropoda." Pp. 259-322 in Weishampel, D.B., Dodson, P. and Osmolska, H. (eds.): The Dinosauria, Second Edition. University of California Press, Berkeley. ISBN 978-0-520-24209-8
- 1 2 3 4 Foster, J. (2007). "Brachiosaurus altithorax." Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. pp. 205–208.
- ↑ Wedel, M.J. (2003). "Vertebral pneumaticity, air sacs, and the physiology of sauropod dinosaurs". Paleobiology. 29 (2): 243–255. doi:10.1666/0094-8373(2003)029<0243:vpasat>2.0.co;2.
- ↑ Wedel, M.J. (2003). "The evolution of vertebral pneumaticity in sauropod dinosaurs". Journal of Vertebrate Paleontology. 23 (2): 344–357. doi:10.1671/0272-4634(2003)023[0344:teovpi]2.0.co;2.
- 1 2 3 4 5 6 7 8 9 10 Riggs, E.S. (1903). "Brachiosaurus altithorax, the largest known dinosaur". American Journal of Science. 4. 15 (88): 299–306.
- 1 2 Glut, D.F. (1997). "Brachiosaurus". Dinosaurs: The Encyclopedia. McFarland & Company. pp. 213–221. ISBN 978-0-89950-917-4.
- 1 2 Turner, C.E.; Peterson, F. (1999). "Biostratigraphy of dinosaurs in the Upper Jurassic Morrison Formation of the Western Interior, USA". In Gillete, David D. Vertebrate Paleontology in Utah. Miscellaneous Publication 99-1. Salt Lake City, Utah: Utah Geological Survey. pp. 77–114. ISBN 978-1-55791-634-1.
- 1 2 Chenoweth, W.L. (1987). "The Riggs Hill and Dinosaur Hill sites, Mesa County, Colorado". In Averett, W. R. Paleontology and Geology of the Dinosaur Triangle. Grand Junction, Colorado: Museum of Western Colorado. pp. 97–100. LCCN 93247073. OCLC 680488874.
- ↑ Lohman, S.W. (1965). Geology and artesian water supply of the Grand Junction area, Colorado. Professional Paper 451. Reston, Virginia: U.S. Geological Survey.
- 1 2 3 4 Riggs, E.S. (1091). "The largest known dinosaur". Science. 13 (327): 549–550. Bibcode:1901Sci....13..549R. PMID 17801098. doi:10.1126/science.13.327.549-a.
- 1 2 3 Riggs, E.S. (1904). "Structure and relationships of opisthocoelian dinosaurs. Part II. The Brachiosauridae". Geological Series (Field Columbian Museum). 2 (6): 229–247.
- ↑ θώραξ, Henry George Liddell, Robert Scott, A Greek-English Lexicon, on Perseus Digital Library
- ↑ Marsh, O.C. (1891). "Restoration of Triceratops" (pdf). American Journal of Science. 41 (244): 339–342. doi:10.2475/ajs.s3-41.244.339.
- ↑ McIntosh, J.S.; Berman, D.S. (1975). "Description of the palate and lower jaw of the sauropod dinosaur Diplodocus (Reptilia: Saurischia) with remarks on the nature of the skull of Apatosaurus". Journal of Paleontology. 49 (1): 187–199.
- 1 2 3 Carpenter, K. and Tidwell, V. (1998). "Preliminary description of a Brachiosaurus skull from Felch Quarry 1, Garden Park, Colorado." Pp. 69–84 in: Carpenter, K., Chure, D. and Kirkland, J. (eds.), The Upper Jurassic Morrison Formation: An Interdisciplinary Study. Modern Geology, 23:1-4.
- ↑ "Expect Awe-Struck Travelers" (Press release). The Field Museum. November 26, 1999. Retrieved 2009-08-27.
- ↑ The Field Museum. "Captions from Selected Historical Photographs (caption number GN89396_52c)." The Field Museum Photo Archives. pdf link. Accessed 2009-Aug-27.
- 1 2 3 Jensen, J.A. (1987). "New brachiosaur material from the Late Jurassic of Utah and Colorado". The Great Basin Naturalist. 47 (4): 592–608.
- 1 2 Curtice, B., Stadtman, K., and Curtice, L. (1996) "A re-assessment of Ultrasauros macintoshi (Jensen, 1985)." Pp. 87-95 in M. Morales (ed.), The Continental Jurassic: Transactions of the Continental Jurassic Symposium, Museum of Northern Arizona Bulletin number 60.
- ↑ Curtice, B.; Stadtman, K. (2001). "The demise of Dystylosaurus edwini and a revision of Supersaurus vivianae". In McCord, R.D.; Boaz, D. Western Association of Vertebrate Paleontologists and Southwest Paleontological Symposium - Proceedings 2001. Mesa Southwest Museum Bulletin. 8. pp. 33–40.
- ↑ Bonnan, M.F.; Wedel, M.J. (2004). "First occurrence of Brachiosaurus (Dinosauria, Sauropoda) from the Upper Jurassic Morrison Formation of Oklahoma". PaleoBios. 24 (2): 12–21.
- ↑ Jensen, J.A. (1985). "Three new sauropod dinosaurs from the Upper Jurassic of Colorado". The Great Basin Naturalist. 45 (4): 697–709.
- 1 2 Olshevsky, G (1991). "A revision of the parainfraclass Archosauria Cope, 1869, excluding the advanced Crocodylia". Mesozoic Meanderings. 2: 1–196.
- 1 2 D'Emic, M. D. (2012). "The early evolution of titanosauriform sauropod dinosaurs". Zoological Journal of the Linnean Society. 166 (3): 624–671. doi:10.1111/j.1096-3642.2012.00853.x.
- ↑ de Lapparent, A.F.; Zbyszewski, G. (1957). "Les dinosauriens du Portugal". Mémoire Service géologique Portugal. 2: 1–63.
- 1 2 Antunes, M.; Mateus, O. (2003). "Dinosaurs of Portugal". Comptes Rendus Palevol. 2 (1): 77–95. doi:10.1016/S1631-0683(03)00003-4.
- 1 2 Janensch, W (1914). "Übersicht über der Wirbeltierfauna der Tendaguru-Schichten nebst einer kurzen Charakterisierung der neu aufgefuhrten Arten von Sauropoden". Archiv für Biontologie. 3: 81–110.
- 1 2 3 Janensch, W. (1929). "Material und Formengehalt der Sauropoden in der Ausbeute der Tendaguru-Expedition." Palaeontographica (Suppl. 7) 2:1–34.
- ↑ Janensch, W. (1935-1936). "Die Schädel der Sauropoden Brachiosaurus, Barosaurus und Dicraeosaurus aus den Tendaguru-Schichten Deutsch-Ostafrikas". Palaeontographica (Suppl. 7) 2: 147-298
- ↑ Taylor, M.P. (2011). "Correction: A re-evaluation of Brachiosaurus altithorax Riggs 1903 (Dinosauria, Sauropoda) and its generic separation from Giraffatitan brancai (Janensch 1914)".". Journal of Vertebrate Paleontology. 31 (3): 727. doi:10.1080/02724634.2011.557115.
- 1 2 3 de Lapparent, A.F. (1960): "Les dinosauriens du "continental intercalaire" du Sahara central" ("The dinosaurs of the "continental intercalaire" of the central Sahara.") Mémoires de la Société Géologic de France, Nouvelle Série 88A vol.39(1-6):1-57. [in French; a translated version, by Matthew Carrano (pdf, no figures), is available through the Polyglot Paleontologist]
- ↑ Mannion, P. D.; Upchurch, P.; Barnes, R. N.; Mateus, O. (2013). "Osteology of the Late Jurassic Portuguese sauropod dinosaur Lusotitan atalaiensis (Macronaria) and the evolutionary history of basal titanosauriforms" (PDF). Zoological Journal of the Linnean Society. 168: 98–206. doi:10.1111/zoj.12029.
- ↑ Carballido, J.L.; Marpmann, J.S.; Schwarz-Wings, D.; Pabst, B. (2012). "New information on a juvenile sauropod specimen from the Morrison Formation and the reassessment of its systematic position". Palaeontology. 55 (2): 567–582. doi:10.1111/j.1475-4983.2012.01139.x.
- ↑ Maier, G. (2003). African dinosaurs unearthed. The Tendaguru Expeditions." Bloomington, IN: Indiana University Press.
- ↑ Taylor, M. (18 Nov. 2009): CT-scanning the Archbishop. Sauropod Vertebrate Picture of the Week (Blog) post at http://svpow.wordpress.com/2009/11/18/ct-scanning-the-archbishop/.
- ↑ Janensch, W. (1950). "Die Wirbelsäule von Brachiosaurus brancai." Palaeontographica (Suppl. 7) 3:27–93.
- ↑ Janensch, W. (1961). "Die Gliedmaßen und Gliedmaßengürtel der Sauropoden der Tendaguru-Schichten." Palaeontographica (Suppl. 7) 3:177–235.
- ↑ Lambert, David; the Diagram Group (1990). "Brachiosaurids". The Dinosaur Data Book. New York: Avon Books. p. 142. ISBN 978-0-380-75896-8.
- ↑ Chure, D.; Britt, B.; Whitlock, J. A.; Wilson, J. A. (2010). "First complete sauropod dinosaur skull from the Cretaceous of the Americas and the evolution of sauropod dentition" (PDF). Naturwissenschaften. 97 (4): 379–391. Bibcode:2010NW.....97..379C. PMC 2841758 . PMID 20179896. doi:10.1007/s00114-010-0650-6.
- 1 2 Ksepka, D. T.; Norell, M. A. (2010). "The illusory evidence for Asian Brachiosauridae: new material of Erketu ellisoni and a phylogenetic appraisal of basal Titanosauriformes" (pdf). American Museum Novitates. 3700: 1–27. doi:10.1206/3700.2.
- 1 2 3 Stevens, K. A.; Parrish, M. J. (1999). "Neck posture and feeding habits of two Jurassic sauropod dinosaurs". Science. 284 (5415): 798–800. Bibcode:1999Sci...284..798S. PMID 10221910. doi:10.1126/science.284.5415.798.
- 1 2 3 Stevens, K. A. and Parrish, M. J. (2005). "Digital reconstructions of sauropod dinosaurs and implications for feeding." In The sauropods: evolution and paleobiology (eds. J. A.Wilson & K. Curry-Rogers), pp. 178–200. Berkeley, CA: University of California Press.
- 1 2 3 Stevens, K. A. and Parrish, M. J. (2005). "Neck posture, dentition and feeding strategies in Jurassic sauropod dinosaurs." In Thunder Lizards: The Sauropodomorph dinosaurs" (eds. V. Tidwell & K. Carpenter). Bloomington, IN: Indiana University Press.
- ↑ Dzemski, G.; Christian, A. (2007). "Flexibility along the neck of the ostrich (Struthio camelus) and consequences for the reconstruction of dinosaurs with extreme neck length". Journal of Morphology. 268 (8): 701–714. doi:10.1002/jmor.10542.
- ↑ Hummel, J.; Gee, C.T.; Südekum, K.-H.; Sander, P.M.; Nogge, G.; Clauss, M. (2008). "In vitro digestibility of fern and gymnosperm foliage: implications for sauropod feeding ecology and diet selection". Proceedings of the Royal Society B. 275 (1638): 1015–1021. PMC 2600911 . PMID 18252667. doi:10.1098/rspb.2007.1728.
- ↑ Barrett, Paul M.; Upchurch, Paul (2005). "Sauropodomorph diversity through time". In Curry Rogers; Kristina A.; Wilson, Jeffrey A. The Sauropods: Evolution and Paleobiology. Berkeley, CA: University of California. pp. 125–156. ISBN 978-0-520-24623-2.
- ↑ Mallison, H. (2011). "Rearing Giants – kinetic-dynamic modeling of sauropod bipedal and tripodal poses." In Klein, N., Remes, K., Gee, C. & Sander M. (eds): Biology of the Sauropod Dinosaurs: Understanding the life of giants. Life of the Past (series ed. Farlow, J.). Bloomington, IN: Indiana University Press.
- 1 2 Sander, P.M.; Christian, A.; Clauss, M.; Fechner, R.; Gee, C.T.; Griebeler, E.-M.; Gunga, H.-C.; Hummel, J.; Mallison, H.; Perry, S.F.; Preuschoft, H.; Rauhut, O.W.M.; Remes, K.; Tütken, T.; Wings, O.; Witzel, U. (2010). "Biology of the sauropod dinosaurs: the evolution of gigantism". Biology Reviews. 86 (1): 117–155. PMC 3045712 . PMID 21251189. doi:10.1111/j.1469-185X.2010.00137.x.
- 1 2 Russell, D. A. (1989). An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press. p. 78. ISBN 978-1-55971-038-1.
- 1 2 Russell, D. A. (1989). An Odyssey in Time: Dinosaurs of North America. Minocqua, Wisconsin: NorthWord Press. pp. 64–70. ISBN 978-1-55971-038-1.
- ↑ Engelmann, G.F.; Chure, D.J.; Fiorillo, A.R. (2004). "The implications of a dry climate for the paleoecology of the fauna of the Upper Jurassic Morrison Formation". Sedimentary Geology. 167 (3–4): 297–308. Bibcode:2004SedG..167..297E. doi:10.1016/j.sedgeo.2004.01.008.
- ↑ Carpenter, K. (2006). "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus". In Foster, J. R.; Lucas, S. G. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 131–138.
- ↑ Chure, D.J.; Litwin, R.; Hasiotis, S.T.; Evanoff, E.; Carpenter, K. (2006). "The fauna and flora of the Morrison Formation: 2006". In Foster, J.R.; Lucas, S.G. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 233–248.
- ↑ Turner, C.E. and Peterson, F., (1999). "Biostratigraphy of dinosaurs in the Upper Jurassic Morrison Formation of the Western Interior, U.S.A." Pp. 77–114 in Gillette, D.D. (ed.), Vertebrate Paleontology in Utah. Utah Geological Survey Miscellaneous Publication 99-1.
- ↑ Foster, J. (2007). "Appendix." Jurassic West: The Dinosaurs of the Morrison Formation and Their World. Indiana University Press. pp. 327–329.
- ↑ Chure, Daniel J.; Litwin, Ron; Hasiotis, Stephen T.; Evanoff, Emmett; Carpenter, Kenneth (2006). "The fauna and flora of the Morrison Formation: 2006". In Foster, John R.; Lucas, Spencer G. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 233–248.
- ↑ Dodson, Peter; Behrensmeyer, A.K.; Bakker, Robert T.; McIntosh, John S. (1980). "Taphonomy and paleoecology of the dinosaur beds of the Jurassic Morrison Formation". Paleobiology. 6 (2): 208–232.
- ↑ Foster, John R. (2003). Paleoecological Analysis of the Vertebrate Fauna of the Morrison Formation (Upper Jurassic), Rocky Mountain Region, U.S.A. New Mexico Museum of Natural History and Science Bulletin, 23. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. p. 29.
- ↑ Mateus, Octávio (2006). "Jurassic dinosaurs from the Morrison Formation (USA), the Lourinhã and Alcobaça Formations (Portugal), and the Tendaguru Beds (Tanzania): A comparison". In Foster, John R.; Lucas, Spencer G. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 223–231.
- ↑ Carpenter, Kenneth (2006). "Biggest of the big: a critical re-evaluation of the mega-sauropod Amphicoelias fragillimus". In Foster, John R.; Lucas, Spencer G. Paleontology and Geology of the Upper Jurassic Morrison Formation. New Mexico Museum of Natural History and Science Bulletin, 36. Albuquerque, New Mexico: New Mexico Museum of Natural History and Science. pp. 131–138.
- ↑ "JPL Small-Body Database Browser: 9954 Brachiosaurus (1991 GX7)". NASA. Retrieved 2007-04-28.
- ↑ Williams, G. "Minor Planet Names: Alphabetical List". Smithsonian Astrophysical Observatory. Retrieved 2007-02-10.
- ↑ "Ronto". Databank. Star Wars.com. Archived from the original on October 3, 2008. Retrieved 2009-01-13.
External links
| Look up Brachiosaurus in Wiktionary, the free dictionary. |
| Wikimedia Commons has media related to Brachiosaurus. |
- Expect awe-struck travelers, from the Field Museum. (O'Hare airport mount)
- Dinosaurier-info, fact-sheet with picture (in German)
- Benes, Josef. Prehistoric Animals and Plants: old-fashioned illustration of Brachiosaurus, bottom left.
- Why was Brachiosaurus that big? Short summary of a peer review paper
- dB Brachiosaurus (Natural History Museum)