Adrenergic receptor
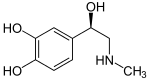

The adrenergic receptors (or adrenoceptors) are a class of G protein-coupled receptors that are targets of the catecholamines, especially norepinephrine (noradrenaline) and epinephrine (adrenaline).
Many cells possess these receptors, and the binding of a catecholamine to the receptor will generally stimulate the sympathetic nervous system. The sympathetic nervous system is responsible for the fight-or-flight response, which includes dilating the pupil, increasing heart rate, mobilizing energy, and diverting blood flow from non-essential organs to skeletal muscle.
History
By the turn of the 19th century, it was agreed that the stimulation of sympathetic nerves could cause different effects on body tissues, depending on the conditions of stimulation (such as the presence or absence of some toxin). Over the first half of the 20th century, two main proposals were made to explain this phenomenon:
- There were (at least) two different types of neurotransmitter released from sympathetic nerve terminals, or
- There were (at least) two different types of detector mechanisms for a single neurotransmitter.
The first hypothesis was championed by Walter Cannon and Arturo Rosenblueth,[1] who interpreted many experiments to then propose that there were two neurotransmitter substances, which they called sympathin E (for 'excitation') and sympathin I (for 'inhibition').
The second hypothesis found support from 1906 to 1913, when Henry Dale explored the effects of adrenaline (which he called adrenine at the time), injected into animals, on blood pressure. Usually, adrenaline would increase the blood pressure of these animals. Although, if the animal had been exposed to ergotoxine, the blood pressure decreased.[2][3] He proposed that the ergotoxine caused "selective paralysis of motor myoneural junctions" (i.e. those tending to increase the blood pressure) hence revealing that under normal conditions that there was a "mixed response", including a mechanism that would relax smooth muscle and cause a fall in blood pressure. This "mixed response", with the same compound causing either contraction or relaxation, was conceived of as the response of different types of junctions to the same compound.
This line of experiments were developed by several groups, including Marsh and colleagues,[4] who in February 1948 showed that a series of compounds structurally related to adrenaline could also show either contracting or relaxing effects, depending on whether or not other toxins were present. This again supported the argument that the muscles had two different mechanisms by which they could respond to the same compound. In June of that year, Raymond Ahlquist, Professor of Pharmacology at Medical College of Georgia, published a paper concerning adrenergic nervous transmission.[5] In it, he explicitly named the different responses as due to what he called α receptors and β receptors, and that the only sympathetic transmitter was adrenaline. While the latter conclusion was subsequently shown to be incorrect (it is now known to be noradrenaline), his receptor nomenclature and concept of two different types of dectors mechanisms for a single neurotransmitter, remains. In 1954, he was able to incorporate his findings in a textbook, Drill's Pharmacology in Medicine,[6] and thereby promulgate the role played by α and β receptor sites in the adrenaline/noradrenaline cellular mechanism. These concepts would revolutionise advances in pharmacotherapeutic research, allowing the selective design of specific molecules to target medical ailments rather than rely upon traditional research into the efficacy of pre-existing herbal medicines.
Categories
There are two main groups of adrenergic receptors, α and β, with several subtypes.
- α receptors have the subtypes α1 (a Gq coupled receptor) and α2 (a Gi coupled receptor[7]). Phenylephrine is a selective agonist of the α receptor.
- β receptors have the subtypes β1, β2 and β3. All three are linked to Gs proteins (although β2 also couples to Gi),[8] which in turn are linked to adenylate cyclase. Agonist binding thus causes a rise in the intracellular concentration of the second messenger cAMP. Downstream effectors of cAMP include cAMP-dependent protein kinase (PKA), which mediates some of the intracellular events following hormone binding. Isoprenaline is a non-selective agonist.
Roles in circulation
Epinephrine (adrenaline) reacts with both α- and β-adrenoreceptors, causing vasoconstriction and vasodilation, respectively. Although α receptors are less sensitive to epinephrine, when activated at pharmacologic doses, they override the vasodilation mediated by β-adrenoreceptors because there are more peripheral α1 receptors than β-adrenoreceptors. The result is that high levels of circulating epinephrine cause vasoconstriction. At lower levels of circulating epinephrine (physiologic epinephrine secretion), β-adrenoreceptor stimulation dominates since epinephrine has a higher affinity for the β2 adrenoreceptor than the α1 adrenoreceptor, producing vasodilation followed by decrease of peripheral vascular resistance.
Subtypes
Smooth muscle behavior is variable depending on anatomical location. Smooth muscle contraction/relaxation is generalized below. One important note is the differential effects of increased cAMP in smooth muscle compared to cardiac muscle. Increased cAMP will promote relaxation in smooth muscle, while promoting increased contractility and pulse rate in cardiac muscle.
†There is no α1C receptor. At one time, there was a subtype known as C, but was found to be identical to one of the previously discovered subtypes. To avoid confusion, naming was continued with the letter D.
α receptors
α receptors have several functions in common, but also individual effects. Common (or still unspecified) effects include:
- Vasoconstriction of veins[10]
- Decrease motility of smooth muscle in gastrointestinal tract[11]
α1 receptor
α1-adrenergic receptors are members of the Gq protein-coupled receptor superfamily. Upon activation, a heterotrimeric G protein, Gq, activates phospholipase C (PLC). The PLC cleaves phosphatidylinositol 4,5-bisphosphate (PIP2), which in turn causes an increase in inositol triphosphate (IP3) and diacylglycerol (DAG). The former interacts with calcium channels of endoplasmic and sarcoplasmic reticulum, thus changing the calcium content in a cell. This triggers all other effects, including a prominent slow after depolarizing current (sADP) in neurons [12]
Specific actions of the α1 receptor mainly involve smooth muscle contraction. It causes vasoconstriction in many blood vessels, including those of the skin, gastrointestinal system, kidney (renal artery)[13] and brain.[14] Other areas of smooth muscle contraction are:
- ureter
- vas deferens
- hair (arrector pili muscles)
- uterus (when pregnant)
- urethral sphincter
- urothelium and lamina propria[15]
- bronchioles (although minor relative to the relaxing effect of β2 receptor on bronchioles)
- blood vessels of ciliary body (stimulation causes mydriasis)
Further effects include glycogenolysis and gluconeogenesis from adipose tissue[16] and liver, as well as secretion from sweat glands[16] and Na+ reabsorption from kidney.[16]
Antagonists may be used primarily in hypertension, anxiety disorder, and panic attacks.
α2 receptor
The α2 receptor couples to the Gi/o protein.[7] It is a presynaptic receptor, causing negative feedback on, for example, norepinephrine (NE). When NE is released into the synapse, it feeds back on the α2 receptor, causing less NE release from the presynaptic neuron. This decreases the effect of NE. There are also α2 receptors on the nerve terminal membrane of the post-synaptic adrenergic neuron.
There are 3 highly homologous subtypes of α2 receptors: α2A, α2Β, and α2C.
Specific actions of the α2 receptor include:
- inhibition of insulin release in the pancreas.[16]
- induction of glucagon release from the pancreas.
- contraction of sphincters of the gastrointestinal tract
- negative feedback in the neuronal synapses - presynaptic inhibition of norepinephrine (NE) release in CNS
- increased thrombocyte aggregation
β receptors
β1 receptor
Specific actions of the β1 receptor include:
- Increase cardiac output by increasing heart rate (positive chronotropic effect), conduction velocity (positive dromotropic effect), and stroke volume (by enhancing contractility—positive inotropic effect).
- Increase renin secretion from juxtaglomerular cells of the kidney.
- Increase ghrelin secretion from the stomach.[17]
β2 receptor
The β2 receptor "binds epinephrine and is involved in the fight or flight response".[18]
Specific actions of the β2 receptor include the following:
- Smooth muscle relaxation, e.g. in bronchi,[16] GI tract (decreased motility), vasodilation of blood vessels, especially those to skeletal muscle (in contrast to vasoconstriction caused by alpha1 and alpha2 adrenoceptors, which is usually the dominant effect).[19]
- Lipolysis in adipose tissue.[20]
- Anabolism in skeletal muscle.[21][22]
- Relax non-pregnant uterus
- Relax detrusor urinae muscle of bladder wall
- Dilate arteries to skeletal muscle
- Glycogenolysis and gluconeogenesis
- Stimulates insulin secretion[23]
- Contract sphincters of GI tract
- Thickened secretions from salivary glands.[16]
- Inhibit histamine-release from mast cells
- Increase renin secretion from kidney[24]
- Relaxation of Bronchioles (salbutamol, a β2 agonist relieves bronchiole constriction)
- Involved in brain - immune communication[25]
β3 receptor
Specific actions of the β3 receptor include:
- Enhancement of lipolysis in adipose tissue. β3 activating drugs could theoretically be used as weight-loss agents, but are limited by the side effect of tremors.
See also
References
- ↑ Cannon WB, Rosenbluth A (31 May 1933). "Studies On Conditions Of Activity In Endocrine Organs XXVI: Sympathin E and Sympathin I". American Journal of Physiology. 104 (3): 557–574.
- ↑ Dale HH (May 1906). "On some physiological actions of ergot". The Journal of Physiology. 34 (3): 163–206. PMC 1465771
 . PMID 16992821. doi:10.1113/jphysiol.1906.sp001148.
. PMID 16992821. doi:10.1113/jphysiol.1906.sp001148. - ↑ Dale HH (Jun 1913). "On the action of ergotoxine; with special reference to the existence of sympathetic vasodilators". The Journal of Physiology. 46 (3): 291–300. PMC 1420444 . PMID 16993202. doi:10.1113/jphysiol.1913.sp001592.
- ↑ Marsh DT, Pelletier MH, Rose CA (Feb 1948). "The comparative pharmacology of the N-alkyl-arterenols". The Journal of Pharmacology and Experimental Therapeutics. 92 (2): 108–20. PMID 18903395.
- ↑ Ahlquist RP (Jun 1948). "A study of the adrenotropic receptors". The American Journal of Physiology. 153 (3): 586–600. PMID 18882199.
- ↑ Drill VA (1954). Pharmacology in medicine: a collaborative textbook. New York: McGraw-Hill.
- 1 2 Qin K, Sethi PR, Lambert NA (Aug 2008). "Abundance and stability of complexes containing inactive G protein-coupled receptors and G proteins". FASEB Journal. 22 (8): 2920–7. PMC 2493464 . PMID 18434433. doi:10.1096/fj.08-105775.
- ↑ Chen-Izu Y, Xiao RP, Izu LT, Cheng H, Kuschel M, Spurgeon H, Lakatta EG (Nov 2000). "G(i)-dependent localization of beta(2)-adrenergic receptor signaling to L-type Ca(2+) channels". Biophysical Journal. 79 (5): 2547–56. Bibcode:2000BpJ....79.2547C. PMC 1301137 . PMID 11053129. doi:10.1016/S0006-3495(00)76495-2.
- ↑ Nisoli E, Tonello C, Landi M, Carruba MO (Jan 1996). "Functional studies of the first selective beta 3-adrenergic receptor antagonist SR 59230A in rat brown adipocytes". Molecular Pharmacology. 49 (1): 7–14. PMID 8569714.
- ↑ Elliott J (Aug 1997). "Alpha-adrenoceptors in equine digital veins: evidence for the presence of both alpha1 and alpha2-receptors mediating vasoconstriction". Journal of Veterinary Pharmacology and Therapeutics. 20 (4): 308–17. PMID 9280371. doi:10.1046/j.1365-2885.1997.00078.x.
- ↑ Sagrada A, Fargeas MJ, Bueno L (Aug 1987). "Involvement of alpha-1 and alpha-2 adrenoceptors in the postlaparotomy intestinal motor disturbances in the rat". Gut. 28 (8): 955–9. PMC 1433140 . PMID 2889649. doi:10.1136/gut.28.8.955.
- ↑ Smith RS, Weitz CJ, Araneda RC (Aug 2009). "Excitatory actions of noradrenaline and metabotropic glutamate receptor activation in granule cells of the accessory olfactory bulb". Journal of Neurophysiology. 102 (2): 1103–14. PMC 2724365 . PMID 19474170. doi:10.1152/jn.91093.2008.
- ↑ Schmitz JM, Graham RM, Sagalowsky A, Pettinger WA (Nov 1981). "Renal alpha-1 and alpha-2 adrenergic receptors: biochemical and pharmacological correlations". The Journal of Pharmacology and Experimental Therapeutics. 219 (2): 400–6. PMID 6270306.
- ↑ Circulation & Lung Physiology I M.A.S.T.E.R. Learning Program, UC Davis School of Medicine
- ↑ Moro C, Tajouri L, Chess-Williams R (Jan 2013). "Adrenoceptor function and expression in bladder urothelium and lamina propria". Urology. 81 (1): 211.e1–7. PMID 23200975. doi:10.1016/j.urology.2012.09.011.
- 1 2 3 4 5 6 Fitzpatrick D, Purves D, Augustine G (2004). "Table 20:2". Neuroscience (Third ed.). Sunderland, Mass: Sinauer. ISBN 0-87893-725-0.
- ↑ Zhao TJ, Sakata I, Li RL, Liang G, Richardson JA, Brown MS, et al. (Sep 2010). "Ghrelin secretion stimulated by {beta}1-adrenergic receptors in cultured ghrelinoma cells and in fasted mice". Proceedings of the National Academy of Sciences of the United States of America. 107 (36): 15868–73. Bibcode:2010PNAS..10715868Z. PMC 2936616 . PMID 20713709. doi:10.1073/pnas.1011116107.
- ↑ Liszewski, Kathy (1 October 2015). "Dissecting the Structure of Membrane Proteins". Genetic Engineering & Biotechnology News (paper). 35 (17): 16.(subscription required)
- ↑ "Adrenergic and Cholinergic Receptors in Blood Vessels". Cardiovascular Physiology. Retrieved 5 May 2015.
- ↑ Large V, Hellström L, Reynisdottir S, et al. (Dec 1997). "Human beta-2 adrenoceptor gene polymorphisms are highly frequent in obesity and associate with altered adipocyte beta-2 adrenoceptor function". The Journal of Clinical Investigation. 100 (12): 3005–13. PMC 508512 . PMID 9399946. doi:10.1172/JCI119854.
- ↑ Kline WO, Panaro FJ, Yang H, Bodine SC (Feb 2007). "Rapamycin inhibits the growth and muscle-sparing effects of clenbuterol". Journal of Applied Physiology. 102 (2): 740–7. PMID 17068216. doi:10.1152/japplphysiol.00873.2006.
- ↑ Kamalakkannan G, Petrilli CM, George I, et al. (Apr 2008). "Clenbuterol increases lean muscle mass but not endurance in patients with chronic heart failure". The Journal of Heart and Lung Transplantation. 27 (4): 457–61. PMID 18374884. doi:10.1016/j.healun.2008.01.013.
- ↑ Santulli, G.; Lombardi, A.; Sorriento, D.; Anastasio, A.; Del Giudice, C.; Formisano, P.; Beguinot, F.; Trimarco, B.; Miele, C.; Iaccarino, G. (2012). "Age-Related Impairment in Insulin Release: The Essential Role of 2-Adrenergic Receptor". Diabetes. 61 (3): 692–701. ISSN 0012-1797. PMC 3282797 . PMID 22315324. doi:10.2337/db11-1027.
- ↑ Kim, Soo Mi; Briggs, Josephine P.; Schnermann, Jurgen (2011). "Convergence of major physiological stimuli for renin release on the Gs-alpha/cyclic adenosine monophosphate signaling pathway". Clinical and Experimental Nephrology. 16 (1): 17–24. ISSN 1342-1751. doi:10.1007/s10157-011-0494-1.
- ↑ Elenkov IJ, Wilder RL, Chrousos GP, et al. (Dec 2000). "The sympathetic nerve--an integrative interface between two supersystems: the brain and the immune system". Pharmacological Reviews. 52 (4): 595–638. PMID 11121511.
Further reading
- Rang HP, Dale MM, Ritter JM, Moore PK (2003). "Chapter 11: Noradrenergic transmission". Pharmacology (5th ed.). Elsevier Churchill Livingstone. ISBN 0-443-07145-4.
- Rang HP, Dale MM, Ritter JM, Flower RJ (2007). "Chapter 11: Noradrenergic transmission". Rang and Dale's Pharmacology (6th ed.). Elsevier Churchill Livingstone. pp. 169–170. ISBN 0-443-06911-5.
External links
- Alpha receptors illustrated
- The Adrenergic Receptors
- "Adrenoceptors". IUPHAR Database of Receptors and Ion Channels. International Union of Basic and Clinical Pharmacology.
- Basic Neurochemistry: α- and β-Adrenergic Receptors
- Brief overview of functions of the β3 receptor
- Theory of receptor activation
- Desensitization of β1 receptors
- UMich Orientation of Proteins in Membranes protein/pdbid-2rh1 - 3D structure of β2 adrenergic receptor in membrane