Genome-wide association study

In genetic epidemiology, a genome-wide association study (GWA study, or GWAS), also known as whole genome association study (WGA study, or WGAS), is an examination of many common genetic variants in different individuals to see if any variant is associated with a trait. GWASs typically focus on associations between single-nucleotide polymorphisms (SNPs) and traits like major diseases.
These studies compare the DNA of participants having varying phenotypes for a particular trait or disease. Participants in a GWAS study may be people with a disease (cases) and similar people without (controls), or they may be people with different phenotypes for a particular trait, for example blood pressure. This approach is known as phenotype-first, in which the participants are classified first by their clinical manifestation(s), as opposed to genotype-first. Each person gives a sample of DNA, from which millions of genetic variants are read using SNP arrays. If one type of the variant (one allele) is more frequent in people with the disease, the variant is said to be associated with the disease. The associated SNPs are then considered to mark a region of the human genome that may influence the risk of disease. In contrast to methods that specifically test one or a few genetic regions, the GWA studies investigate the entire genome. The approach is therefore said to be non-candidate-driven in contrast to gene-specific candidate-driven studies. GWA studies identify SNPs and other variants in DNA associated with a disease, but that cannot on their own specify which genes are causal.[2][3][4]
The first successful GWAS was published in 2005. It investigated patients with age-related macular degeneration and found two SNPs with significantly altered allele frequency compared to healthy controls.[5] As of 2011, hundreds or thousands of individuals are tested, over 1,200 human GWA studies have examined over 200 diseases and traits, and almost 4,000 SNP associations have been found.[6] Several GWA studies have received criticism for omitting important quality control steps, rendering the findings invalid, but modern publications address these issues. However, the methodology itself still has opponents.
Background
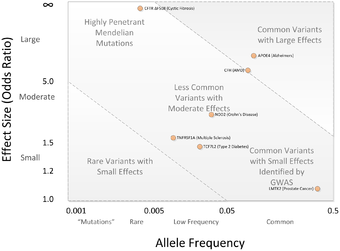
Any two human genomes differ in millions of different ways. There are small variations in the individual nucleotides of the genomes (SNPs) as well as many larger variations, such as deletions, insertions and copy number variations. Any of these may cause alterations in an individual's traits, or phenotype, which can be anything from disease risk to physical properties such as height.[8] Around the year 2000, prior to the introduction of GWA studies, the primary method of investigation was through inheritance studies of genetic linkage in families. This approach had proven highly useful towards single gene disorders.[9] However, for common and complex diseases the results of genetic linkage studies proved hard to reproduce.[8][10] A suggested alternative to linkage studies was the genetic association study. This study type asks if the allele of a genetic variant is found more often than expected in individuals with the phenotype of interest (e.g. with the disease being studied). Early calculations on statistical power indicated that this approach could be better than linkage studies at detecting weak genetic effects.[11]
In addition to the conceptual framework several additional factors enabled the GWA studies. One was the advent of biobanks, which are repositories of human genetic material that greatly reduced the cost and difficulty of collecting sufficient numbers of biological specimens for study.[12] Another was the International HapMap Project, which, from 2003 identified a majority of the common SNPs interrogated in a GWA study.[13] The haploblock structure identified by HapMap project also allowed the focus on the subset of SNPs that would describe most of the variation. Also the development of the methods to genotype all these SNPs using genotyping arrays was an important prerequisite.[14]
Methods

The most common approach of GWA studies is the case-control setup, which compares two large groups of individuals, one healthy control group and one case group affected by a disease. All individuals in each group are genotyped for the majority of common known SNPs. The exact number of SNPs depends on the genotyping technology, but are typically one million or more.[7] For each of these SNPs it is then investigated if the allele frequency is significantly altered between the case and the control group.[16] In such setups, the fundamental unit for reporting effect sizes is the odds ratio. The odds ratio is the ratio of two odds, which in the context of GWA studies are the odds of disease for individuals having a specific allele and the odds of disease for individuals who do not have that same allele. When the allele frequency in the case group is much higher than in the control group, the odds ratio is higher than 1, and vice versa for lower allele frequency. Additionally, a P-value for the significance of the odds ratio is typically calculated using a simple chi-squared test. Finding odds ratios that are significantly different from 1 is the objective of the GWA study because this shows that a SNP is associated with disease.[16]
There are several variations to this case-control approach. A common alternative to case-control GWA studies is the analysis of quantitative phenotypic data, e.g. height or biomarker concentrations or even gene expression. Likewise, alternative statistics designed for dominance or recessive penetrance patterns can be used.[16] Calculations are typically done using bioinformatics software such as SNPTEST and PLINK, which also include support for many of these alternative statistics.[17][18] Earlier GWAS focused on the effect of individual SNPs. However, the empirical evidence shows that complex interactions among two or more SNPs, epistasis, might contribute to complex diseases. Moreover, the researchers tries to integrate GWA data with other biological data such as protein protein interaction network to extract more informative results.[19][20]
A key step in the majority of GWA studies is the imputation of genotypes at SNPs not on the genotype chip used in the study.[21] This process greatly increases the number of SNPs that can be tested for association, increases the power of the study, and facilitates meta-analysis of GWAS across distinct cohorts. Genotype imputation is carried out by statistical methods that combine the GWAS data together with a reference panel of haplotypes. These methods take advantage of sharing of haplotypes between individuals over short stretches of sequence to impute alleles. Existing software packages for genotype imputation include IMPUTE2[22] and MaCH.[23]
In addition to the calculation of association, it is common to take into account any variables that could potentially confound the results. Sex and age are common examples of confounding variables. Moreover, it is also known that many genetic variations are associated with the geographical and historical populations in which the mutations first arose.[24] Because of this association, studies must take account of the geographical and ethnical background of participants by controlling for what is called population stratification.
After odds ratios and P-values have been calculated for all SNPs, a common approach is to create a Manhattan plot. In the context of GWA studies, this plot shows the negative logarithm of the P-value as a function of genomic location. Thus the SNPs with the most significant association stands out on the plot, usually as stacks of points because of haploblock structure. Importantly, the P-value threshold for significance is corrected for multiple testing issues. The exact threshold varies by study,[25] but the conventional threshold is 5×10−8 to be significant in the face of hundreds of thousands to millions of tested SNPs.[7][16][26] GWA studies typically perform the first analysis in a discovery cohort, followed by validation of the most significant SNPs in an independent validation cohort.
Results

Attempts have been made at creating comprehensive catalogues of SNPs that have been identified from GWA studies.[28] As of 2009, SNPs associated with diseases are numbered in the thousands.[6]
The first GWA study, conducted in 2005, compared 96 patients with age-related macular degeneration (ARMD) with 50 healthy controls.[29] It identified two SNPs with significantly altered allele frequency between the two groups. These SNPs were located in the gene encoding complement factor H, which was an unexpected finding in the research of ARMD. The findings from these first GWA studies have subsequently prompted further functional research towards therapeutical manipulation of the complement system in ARMD.[30] Another landmark publication in the history of GWA studies was the Wellcome Trust Case Control Consortium (WTCCC) study, the largest GWA study ever conducted at the time of its publication in 2007. The WTCCC included 14,000 cases of seven common diseases (~2,000 individuals for each of coronary heart disease, type 1 diabetes, type 2 diabetes, rheumatoid arthritis, Crohn's disease, bipolar disorder, and hypertension) and 3,000 shared controls.[31] This study was successful in uncovering many new disease genes underlying these diseases.[31][32]
Since these first landmark GWA studies, there have been two general trends.[33] One has been towards larger and larger sample sizes. At the end of 2011, the largest sample sizes were in the range of 200,000 individuals.[34] The reason is the drive towards reliably detecting risk-SNPs that have smaller odds ratios and lower allele frequency. Another trend has been towards the use of more narrowly defined phenotypes, such as blood lipids, proinsulin or similar biomarkers.[35][36] These are called intermediate phenotypes, and their analyses may be of value to functional research into biomarkers.[37]
A central point of debate on GWA studies has been that most of the SNP variations found by GWA studies are associated with only a small increased risk of the disease, and have only a small predictive value. The median odds ratio is 1.33 per risk-SNP, with only a few showing odds ratios above 3.0.[2][38] These magnitudes are considered small because they do not explain much of the heritable variation. This heritable variation is known from heritability studies based on monozygotic twins.[39] For example, it is known that 80–90% of height is heritable. Of these 80-90%, however, the GWA studies only account for a minority.[39]
Clinical applications
A challenge for future successful GWA study is to apply the findings in a way that accelerates drug and diagnostics development, including better integration of genetic studies into the drug-development process and a focus on the role of genetic variation in maintaining health as a blueprint for designing new drugs and diagnostics.[40] Several studies have looked into the use of risk-SNP markers as a means of directly improving the accuracy of prognosis. Some have found that the accuracy of prognosis improves,[41] while others report only minor benefits from this use.[42] Generally, a problem with this direct approach is the small magnitudes of the effects observed. A small effect ultimately translates into a poor separation of cases and controls and thus only a small improvement of prognosis accuracy. An alternative application is therefore the potential for GWA studies to elucidate pathophysiology.[43]
One such success is related to identifying the genetic variant associated with response to anti-hepatitis C virus treatment. For genotype 1 hepatitis C treated with Pegylated interferon-alpha-2a or Pegylated interferon-alpha-2b combined with ribavirin, a GWA study[44] has shown that SNPs near the human IL28B gene, encoding interferon lambda 3, are associated with significant differences in response to the treatment. A later report demonstrated that the same genetic variants are also associated with the natural clearance of the genotype 1 hepatitis C virus.[45] These major findings facilitated the development of personalized medicine and allowed physicians to customize medical decisions based on the patient's genotype.[46]
The goal of elucidating pathophysiology has also led to increased interest in the association between risk-SNPs and the gene expression of nearby genes, the so-called expression quantitative trait loci (eQTL) studies.[47] The reason is that GWAS studies identify risk-SNPs, but not risk-genes, and specification of genes is one step closer towards actionable drug targets.[48] As a result, major GWA studies of 2011 typically included extensive eQTL analysis.[49][50][51] One of the strongest eQTL effects observed for a GWA-identified risk SNP is the SORT1 locus.[35] Functional follow up studies of this locus using small interfering RNA and gene knock-out mice have shed light on the metabolism of low-density lipoproteins, which have important clinical implications for cardiovascular disease.[35][52][53]
Limitations
GWA studies have several issues and limitations that can be taken care of through proper quality control and study setup. Lack of well defined case and control groups, insufficient sample size, control for multiple testing and control for population stratification are common problems.[3] Given the large sample sizes typical of GWA studies, statistical issues associated with multiple testing correction may be problematic, to this end it has been noted "the GWA approach can be problematic because the massive number of statistical tests performed presents an unprecedented potential for false-positive results".[3] Ignoring these correctible issues has been cited as contributing to a general sense of problems with the GWA methodology.[54] In addition to easily correctible problems such as these, some more subtle but important issues have surfaced. A high profile GWA study that investigated individuals with very long life spans to identify SNPs associated with longevity is an example of this.[55] The publication came under scrutiny because of a discrepancy between the type of genotyping array in the case and control group, which caused several SNPs to be falsely highlighted as associated with longevity.[56] The study was subsequently retracted.[57]
In addition to these preventable issues, GWA studies have attracted more fundamental criticism, mainly because of their assumption that common genetic variation plays a large role in explaining the heritable variation of common disease.[58] This aspect of GWA studies has attracted the criticism that, although it could not have been known prospectively, GWA studies were ultimately not worth the expenditure.[43] Alternative strategies suggested involve linkage analysis. More recently, the rapidly decreasing price of complete genome sequencing have also provided a realistic alternative to genotyping array-based GWA studies. It can be discussed if the use of this new technique is still referred to as a GWA study, but high-throughput sequencing does have potential to side-step some of the shortcomings of non-sequencing GWA.[59]
See also
- Association mapping
- Epidemiology
- Gene-environment interaction
- Genomics
- Linkage disequilibrium
- Molecular epidemiology
References
- ↑ Ikram MK; Sim X; Xueling S; et al. (October 2010). McCarthy, Mark I, ed. "Four novel Loci (19q13, 6q24, 12q24, and 5q14) influence the microcirculation in vivo". PLoS Genet. 6 (10): e1001184. doi:10.1371/journal.pgen.1001184. PMC 2965750. PMID 21060863.
- 1 2 Manolio TA; Guttmacher, Alan E.; Manolio, Teri A. (July 2010). "Genomewide association studies and assessment of the risk of disease". N. Engl. J. Med. 363 (2): 166–76. doi:10.1056/NEJMra0905980. PMID 20647212.
- 1 2 3 Pearson TA; Manolio TA (March 2008). "How to interpret a genome-wide association study". JAMA 299 (11): 1335–44. doi:10.1001/jama.299.11.1335. PMID 18349094.
- ↑ "Genome-Wide Association Studies". National Human Genome Research Institute.
- ↑ Klein RJ; Zeiss C; Chew EY; Tsai JY; et al. (April 2005). "Complement Factor H Polymorphism in Age-Related Macular Degeneration". Science 308 (5720): 385–9. doi:10.1126/science.1109557. PMC 1512523. PMID 15761122.
- 1 2 Johnson AD; O'Donnell CJ (2009). "An Open Access Database of Genome-wide Association Results". BMC Med. Genet. 10: 6. doi:10.1186/1471-2350-10-6. PMC 2639349. PMID 19161620.
- 1 2 3 Bush WS; Moore JH (2012). Lewitter, Fran; Kann, Maricel, eds. "Chapter 11: genome-wide association studies". PLoS Comput Biol 8 (12): e1002822. doi:10.1371/journal.pcbi.1002822. PMC 3531285. PMID 23300413.
- 1 2 Strachan T; Read A. Human Molecular Genetics (4th ed.). Garland Science. pp. 467–495. ISBN 978-0-8153-4149-9.
- ↑ "Online Mendelian Inheritance in Man". Retrieved 2011-12-06.
- ↑ Altmüller J; Palmer LJ; Fischer G; Scherb H; et al. (November 2001). "Genomewide Scans of Complex Human Diseases: True Linkage Is Hard to Find". Am. J. Hum. Genet. 69 (5): 936–50. doi:10.1086/324069. PMC 1274370. PMID 11565063.
- ↑ Risch N; Merikangas K (September 1996). "The future of genetic studies of complex human diseases". Science 273 (5281): 1516–7. doi:10.1126/science.273.5281.1516. PMID 8801636.
- ↑ Greely HT (2007). "The uneasy ethical and legal underpinnings of large-scale genomic biobanks". Annu Rev Genomics Hum Genet 8: 343–64. doi:10.1146/annurev.genom.7.080505.115721. PMID 17550341.
- ↑ The International HapMap Project, Gibbs RA, Belmont JW, Hardenbol P, Willis TD, Yu F, Yang H, Ch'Ang L-Y, Huang W (December 2003). "The International HapMap Project". Nature 426 (6968): 789–96. doi:10.1038/nature02168. PMID 14685227.
- ↑ Schena M; Shalon D; Davis RW; Brown PO (October 1995). "Quantitative monitoring of gene expression patterns with a complementary DNA microarray". Science 270 (5235): 467–70. doi:10.1126/science.270.5235.467. PMID 7569999.
- ↑ Wellcome Trust Case Control Consortium (June 2007). "Genome-wide association study of 14,000 cases of seven common diseases and 3,000 shared controls". Nature 447 (7145): 661–78. doi:10.1038/nature05911. PMC 2719288. PMID 17554300.
- 1 2 3 4 Clarke GM; Anderson CA; Pettersson FH; Cardon LR; et al. (February 2011). "Basic statistical analysis in genetic case-control studies". Nat Protoc 6 (2): 121–33. doi:10.1038/nprot.2010.182. PMC 3154648. PMID 21293453.
- ↑ "Genome-wide association study of 14,000 cases of seven common diseases and 3,000 shared controls". Nature 447 (7145): 661–678. 2007. doi:10.1038/nature05911. PMC 2719288. PMID 17554300.
- ↑ Purcell S; Neale B; Todd-Brown K; Thomas L; et al. (September 2007). "PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses". Am. J. Hum. Genet. 81 (3): 559–75. doi:10.1086/519795. PMC 1950838. PMID 17701901.
- ↑ Ayati, Marzieh; Erten, Sinan; Chance, Mark R.; Koyutürk, Mehmet (2015-06-30). "MOBAS: identification of disease-associated protein subnetworks using modularity-based scoring". EURASIP Journal on Bioinformatics and Systems Biology 2015 (1): 1–14. doi:10.1186/s13637-015-0025-6. ISSN 1687-4153.
- ↑ Ayati, Marzieh; Koyutürk, Mehmet (2015-01-01). "Assessing the Collective Disease Association of Multiple Genomic Loci". Proceedings of the 6th ACM Conference on Bioinformatics, Computational Biology and Health Informatics. BCB '15 (New York, NY, USA: ACM): 376–385. doi:10.1145/2808719.2808758. ISBN 978-1-4503-3853-0.
- ↑ Marchini J; Howie B (2010). "Genotype imputation for genome-wide association studies". Nature Reviews Genetics 11 (7): 499–511. doi:10.1038/nrg2796. PMID 20517342.
- ↑ Howie B; Marchini J; Stephens M (2011). "Genotype imputation with thousands of genomes". G3: Genes|Genomes|Genetics 1 (6): 457–70. doi:10.1534/g3.111.001198. PMC 3276165. PMID 22384356.
- ↑ Cáp J; Misíková Z (1975). "Chronic myelogenous leukaemia as a possible consequence of immunosuppressive treatment of nephrotic syndrome (author's transl)". Monatsschrift fur Kinderheilkunde 123 (10): 718–20. PMID 1058334.
- ↑ Novembre J; Johnson T; Bryc K; Kutalik Z; et al. (November 2008). "Genes mirror geography within Europe". Nature 456 (7218): 98–101. doi:10.1038/nature07331. PMC 2735096. PMID 18758442.
- ↑ Wittkowski KM; Sonakya V; Bigio B; Tonn MK; et al. (January 2014). "A novel computational biostatistics approach implies impaired dephosphorylation of growth factor receptors as associated with severity of autism". Transl Psychiatry 4 (1): e354. doi:10.1038/tp.2013.124. PMC 3905234. PMID 24473445.
- ↑ Barsh GS; Copenhaver GP; Gibson G; Williams SM (5 July 2012). "Guidelines for Genome-Wide Association Studies". PLoS Genetics 8 (7): e1002812. doi:10.1371/journal.pgen.1002812. PMID 22792080.
- ↑ Sanna S, Li B; Mulas A; Sidore C; Kang HM; et al. (July 2011). Gibson, Greg, ed. "Fine mapping of five loci associated with low-density lipoprotein cholesterol detects variants that double the explained heritability". PLoS Genet. 7 (7): e1002198. doi:10.1371/journal.pgen.1002198. PMC 3145627. PMID 21829380.
- ↑ Hindorff LA; Sethupathy P; Junkins HA; Ramos EM; et al. (June 2009). "Potential etiologic and functional implications of genome-wide association loci for human diseases and traits". Proc. Natl. Acad. Sci. U.S.A. 106 (23): 9362–7. doi:10.1073/pnas.0903103106. PMC 2687147. PMID 19474294.
- ↑ Haines JL; Hauser MA; Schmidt S; Scott WK; et al. (2005). "Complement Factor H Variant Increases the Risk of Age-Related Macular Degeneration". Science 308 (5720): 419–421. doi:10.1126/science.1110359. PMID 15761120.
- ↑ Fridkis-Hareli M; Storek M; Mazsaroff I; Risitano AM; et al. (October 2011). "Design and development of TT30, a novel C3d-targeted C3/C5 convertase inhibitor for treatment of human complement alternative pathway–mediated diseases". Blood 118 (17): 4705–13. doi:10.1182/blood-2011-06-359646. PMC 3208285. PMID 21860027.
- 1 2 Wellcome Trust Case Control Consortium, Burton PR; Clayton DG; Cardon LR; et al. (June 2007). "Genome-wide association study of 14,000 cases of seven common diseases and 3,000 shared controls". Nature 447 (7145): 661–78. doi:10.1038/nature05911. PMC 2719288. PMID 17554300.
- ↑ "Largest ever study of genetics of common diseases published today" (Press release). Wellcome Trust Case Control Consortium. 2007-06-06. Retrieved 2008-06-19.
- ↑ Ioannidis JP; Thomas G; Daly MJ (2009). "Validating, augmenting and refining genome-wide association signals". Nat Rev Genet 10 (5): 318–29. doi:10.1038/nrg2544. PMID 19373277.
- ↑ Ehret GB; Munroe PB; Rice KM; Bochud M; et al. (October 2011). "Genetic variants in novel pathways influence blood pressure and cardiovascular disease risk". Nature 478 (7367): 103–9. doi:10.1038/nature10405. PMC 3340926. PMID 21909115.
- 1 2 3 Kathiresan S; Willer CJ; Peloso GM; Demissie S; et al. (January 2009). "Common variants at 30 loci contribute to polygenic dyslipidemia". Nat. Genet. 41 (1): 56–65. doi:10.1038/ng.291. PMC 2881676. PMID 19060906.
- ↑ Strawbridge RJ; Dupuis J; Prokopenko I; Barker A; et al. (October 2011). "Genome-Wide Association Identifies Nine Common Variants Associated With Fasting Proinsulin Levels and Provides New Insights Into the Pathophysiology of Type 2 Diabetes". Diabetes 60 (10): 2624–34. doi:10.2337/db11-0415. PMC 3178302. PMID 21873549.
- ↑ Danesh J; Pepys MB (November 2009). "C-reactive protein and coronary disease: is there a causal link?". Circulation 120 (21): 2036–9. doi:10.1161/CIRCULATIONAHA.109.907212. PMID 19901186.
- ↑ Ku CS; Loy EY; Pawitan Y; Chia KS (April 2010). "The pursuit of genome-wide association studies: where are we now?". J. Hum. Genet. 55 (4): 195–206. doi:10.1038/jhg.2010.19. PMID 20300123.
- 1 2 Maher B (November 2008). "Personal genomes: The case of the missing heritability". Nature 456 (7218): 18–21. doi:10.1038/456018a. PMID 18987709.
- ↑ Iadonato SP; Katze MG (September 2009). "Genomics: Hepatitis C virus gets personal". Nature 461 (7262): 357–8. doi:10.1038/461357a. PMID 19759611.

- ↑ Muehlschlegel JD; Liu KY; Perry TE; Fox AA; et al. (September 2010). "Chromosome 9p21 Variant Predicts Mortality after CABG Surgery". Circulation 122 (11 Suppl): S60–5. doi:10.1161/CIRCULATIONAHA.109.924233. PMC 2943860. PMID 20837927.
- ↑ Paynter NP; Chasman DI; Paré G; Buring JE; et al. (February 2010). "Association between a Literature-Based Genetic Risk Score and Cardiovascular Events in 19,313 Women". JAMA 303 (7): 631–7. doi:10.1001/jama.2010.119. PMC 2845522. PMID 20159871.
- 1 2 Couzin-Frankel J (June 2010). "Major heart disease genes prove elusive". Science 328 (5983): 1220–1. doi:10.1126/science.328.5983.1220. PMID 20522751.
- ↑ Ge D; Fellay J; Thompson AJ; Simon JS; et al. (September 2009). "Genetic variation in IL28B predicts hepatitis C treatment-induced viral clearance". Nature 461 (7262): 399–401. doi:10.1038/nature08309. PMID 19684573.
- ↑ Thomas DL; Thio CL; Martin MP, Qi Y; Ge D; et al. (October 2009). "Genetic variation in IL28B and spontaneous clearance of hepatitis C virus". Nature 461 (7265): 798–801. doi:10.1038/nature08463. PMC 3172006. PMID 19759533.
- ↑ Lu, YF; Goldstein, DB; Angrist, M; Cavalleri, G (24 July 2014). "Personalized medicine and human genetic diversity". Cold Spring Harbor perspectives in medicine 4 (9): a008581. doi:10.1101/cshperspect.a008581. PMID 25059740.
- ↑ Folkersen L; van't Hooft F; Chernogubova E; Agardh HE; et al. (August 2010). "Association of genetic risk variants with expression of proximal genes identifies novel susceptibility genes for cardiovascular disease". Circ Cardiovasc Genet 3 (4): 365–73. doi:10.1161/CIRCGENETICS.110.948935. PMID 20562444.
- ↑ Fareed M., Afzal M (2013). "Single nucleotide polymorphism in genome-wide association of human population: A tool for broad spectrum service". Egypt. J. Med. Hum. Genet 14: 123–134. doi:10.1016/j.ejmhg.2012.08.001.
- ↑ Bown MJ; Jones GT; Harrison SC; Wright BJ; et al. (November 2011). "Abdominal Aortic Aneurysm Is Associated with a Variant in Low-Density Lipoprotein Receptor-Related Protein 1". Am. J. Hum. Genet. 89 (5): 619–27. doi:10.1016/j.ajhg.2011.10.002. PMC 3213391. PMID 22055160.
- ↑ "A genome-wide association study in Europeans and South Asians identifies five new loci for coronary artery disease". Nat. Genet. 43 (4): 339–44. April 2011. doi:10.1038/ng.782. PMID 21378988.
- ↑ Johnson T; Gaunt TR; Newhouse SJ; Padmanabhan S; et al. (November 2011). "Blood Pressure Loci Identified with a Gene-Centric Array". Am J Hum Genet 89 (6): 688–700. doi:10.1016/j.ajhg.2011.10.013. PMC 3234370. PMID 22100073.
- ↑ Dubé JB; Johansen CT; Hegele RA (June 2011). "Sortilin: an unusual suspect in cholesterol metabolism: from GWAS identification to in vivo biochemical analyses, sortilin has been identified as a novel mediator of human lipoprotein metabolism". BioEssays 33 (6): 430–7. doi:10.1002/bies.201100003. PMID 21462369.
- ↑ Bauer RC; Stylianou IM; Rader DJ (April 2011). "Functional validation of new pathways in lipoprotein metabolism identified by human genetics". Curr. Opin. Lipidol. 22 (2): 123–8. doi:10.1097/MOL.0b013e32834469b3. PMID 21311327.
- ↑ "Size matters, and other lessons from medical genetics". Retrieved 2011-12-07.
- ↑ Sebastiani P; Solovieff N; Puca A; Hartley SW; et al. (July 2010). "Genetic signatures of exceptional longevity in humans". Science 2010. doi:10.1126/science.1190532. PMID 20595579. (Retracted)
- ↑ MacArthur, Daniel. "Serious flaws revealed in "longevity genes" study". Wired. Retrieved 2011-12-07.
- ↑ Sebastiani P; Solovieff N; Puca A; Hartley SW; et al. (July 2011). "Retraction". Science 333 (6041): 404. doi:10.1126/science.333.6041.404-a. PMID 21778381.
- ↑ Visscher PM; Brown MA; McCarthy MI; Yang J (2012). "Five years of GWAS discovery". Am J Hum Genet 90 (1): 7–24. doi:10.1016/j.ajhg.2011.11.029. PMC 3257326. PMID 22243964.
- ↑ Visscher PM; Goddard ME; Derks EM; Wray NR (June 2011). "Evidence-based psychiatric genetics, AKA the false dichotomy between common and rare variant hypotheses". Mol Psychiatry 17 (5): 474–85. doi:10.1038/mp.2011.65. PMID 21670730.
External links
- Genotype-phenotype interaction software tools and databases on Omictools
- Statistical Methods for the Analysis of Genome-Wide Association Studies [video lecture series]
- Whole genome association studies — by the National Human Genome Research Institute
- GWAS Central — a central database of summary-level genetic association findings
- Barrett, Jeff (18 July 2010). "How to read a genome-wide association study". Genomes Unzipped.
- Consortia of genome-wide association studies (GWAS) — by Bennett SN, Caporaso, NE, et al.
- PLINK — whole genome association analysis toolset
- ENCODE threads explorer Impact of functional information on understanding variation. Nature (journal)
- Custom Genome-Wide Association Studies
| ||||||||||||||||||||||