Therocephalia
| Therocephalia Temporal range: Middle Permian–Middle Triassic, 265–243 Ma | |
|---|---|
 | |
| Skull of Ictidosuchoides intermedium in the Wrexham Museum, Wales | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Clade: | Synapsida |
| Order: | Therapsida |
| Clade: | Eutheriodontia |
| Suborder: | †Therocephalia Broom, 1905 |
| Families | |
|
See below | |
Therocephalia is an extinct suborder of eutheriodont therapsids from the Permian and Triassic. The therocephalians ("beast-heads") are named after their large skulls, which, along with the structure of their teeth, suggest that they were carnivores. Like other non-mammalian synapsids, therocephalians are described as mammal-like reptiles. Therocephalia is the group most closely related to the cynodonts, which gave rise to the mammals. This relationship takes evidence in a variety of skeletal features. The phylogeny of therocephalians has been disputed, as the monophyly of the group and the relationships of its members are unclear.
The fossils of therocephalians are numerous in the Karoo of South Africa, but have also been found in Russia, China, and Antarctica. Early therocephalian fossils discovered in Middle Permian deposits of South Africa support a Gondwanan origin for the group, which seems to have spread quickly throughout the world. Although almost every therocephalian lineage ended during the great Permian–Triassic extinction event, a few representatives of the subgroup called Eutherocephalia survived into the Early Triassic. However, the last therocephalians became extinct by the early Middle Triassic, possibly due to climate change and competition with cynodonts and various groups of reptiles.
Anatomy and physiology

Like the gorgonopsids and many cynodonts, most therocephalians were presumably carnivores. The earlier therocephalians were in many respects as primitive as the gorgonopsids, but they did show certain advanced features. There is an enlargement of the temporal opening for broader jaw adductor muscle attachment and a reduction of the phalanges (finger and toe bones) to the mammalian phalangeal formula. The presence of an incipient secondary palate in advanced therocephalians is another feature shared with mammals. The discovery of maxilloturbinal ridges in forms such as the primitive therocephalian Glanosuchus, suggests that at least some therocephalians may have been warm-blooded.
The later therocephalians included the advanced Baurioidea, which carried some theriodont characteristics to a high degree of specialization. For instance, small baurioids and the herbivorous Bauria did not have an ossified postorbital bar separating the orbit from the temporal opening—a condition typical of primitive mammals. These and other advanced features led to the long-held opinion, now rejected, that the ictidosaurs and even some early mammals arose from a baurioid therocephalian stem. Mammalian characteristics such as this seem to have evolved in parallel among a number of different therapsid groups, even within Therocephalia. Several more specialized lifestyles have been suggested for some therocephalians. Many small forms like ictidosuchids have been interpreted as aquatic animals. Evidence for aquatic lifestyles includes sclerotic rings that may have stabilized the eye under the pressure of water and strongly developed cranial joints, which may have supported the skull when consuming large fish and aquatic invertebrates. One therocephalian, Nothogomphodon, had large saber-like canine teeth and may have fed on large animals, including other therocephalians. Other therocephalians such as bauriids and nanictidopids have wide teeth with many ridges similar to those of mammals, and may have been herbivores.[1]
Many small therocephalians have small pits on their snouts that probably supported vibrissae or whiskers. In 1994, Russian paleontologist Leonid Tatarinov proposed that these pits were part of an electroreception system in aquatic therocephalians.[2] However, it is more likely these pits are enlarged versions of the ones thought to support whiskers, or holes for blood vessels in a fleshy lip.[1]
Classification
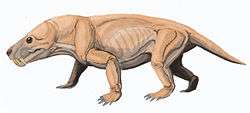
The therocephalians evolved from an early line of pre-mammalian eutheriodont therapsids, and are the sister group to the cynodonts which include mammals and their ancestors. Therocephalians are at least as ancient as a third large branch of therapsids, the gorgonopsids, which they resemble in many primitive features. For example, many early therocephalians possess long canine teeth similar to those of gorgonopsids. The therocephalians, however, outlasted the gorgonopsids, persisting into the early-Middle Triassic period.
While common ancestry with cynodonts (and, thus, mammals) accounts for many similarities among these groups, some scientists believe that other similarities may be better attributed to convergent evolution, such as the loss of the postorbital bar in some forms, a mammalian phalangeal formula, and some form of a secondary palate in most taxa (see below). Therocephalians and cynodonts both survived the Permian-Triassic mass extinction, but while therocephalians soon went extinct, cynodonts underwent rapid diversification. Therocephalians experienced a decreased rate of cladogenesis, meaning that few new groups appeared after the extinction. Most Triassic therocephalian lineages originated in the Late Permian, and lasted for only a short period of time in the Triassic.[3]
Taxonomy



Some previously recognized therocephalian clades have turned out to be artificial. For example, the Scaloposauridae were classified based on fossils with mostly juvenile characteristics, but probably represent immature specimens from other known therocephalian families.
On the other hand, the aberrant therocephalian family Lycosuchidae, once identified by the presence of multiple caniniform teeth, was thought to represent an unnatural group based on a study of canine replacement in that group (van den Heever, 1980). However, subsequent analysis has exposed additional synapomorphies supporting the monophyly of this group, and Lycosuchidae is currently considered the most basal clade within a monophyletic Therocephalia (van den Heever, 1994).
Order Therapsida
- Suborder Therocephalia
- Crapartinella
- Theriodesmus
- Family Lycosuchidae
- Clade Scylacosauria
- Family Scylacosauridae
- Family Trochosuchidae
- Infraorder Eutherocephalia
Phylogeny
Below is a cladogram modified from Sidor (2001) and Huttenlocker (2009):[4][5]
| Therapsida |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
Below is a cladogram modified from analysis published by Adam K. Huttenlocker in 2014.[6] It is based on the data matrix published by Sigurdsen et al. (2012),[7] which is itself a modified version of Huttenlocker et al. (2011).[3] Six additional characters and 22 new ingroup taxa were added to the matrix of Sigurdsen et al. (2012), resulting in a matrix that includes 58 therapsids and outgroup taxa, including 49 therocephalians, which are scored based on 135 morphological traits. Huttenlocker (2014) used this cladogram, among others, to construct an informal supertree for evaluating temporal and phylogenetic distributions of body size in Permo-Triassic eutheriodonts.[6]
| Therapsida |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
In popular culture
- A pack of unspecified therocephalians appeared in the third episode of the BBC series, Walking with Monsters (which look similar to the Thrinaxodons from Walking with Dinosaurs). One of them attacks a Lystrosaurus during the night, injecting the prey with a neurotoxin delivered by a venom gland in the cheek. Although the existence of such a gland is controversial, the absence of postcanine teeth in association with a maxillary pit and grooved caniniform teeth indicate that a venom delivery apparatus may have existed in some therocephalian therapsids (e.g., Euchambersia).
- Also, a large pack of therocephalians appeared in Episode 4.4 of the ITV science-fiction drama Primeval. They were also portrayed as being venomous.
See also
References
- 1 2 Ivakhnenko, M.F. (2011). "Permian and Triassic therocephals (Eutherapsida) of Eastern Europe". Paleontological Journal 45 (9): 981–1144. doi:10.1134/S0031030111090012.
- ↑ Tatarinov, L.P. (1994). "On the preservation of rudimentary rostral tubular complex of crossopterygians in theriodonts and on possible development of the electroreceptor systems in some members of this group". Doklady Akademii Nauk 338 (2): 278–281.
- 1 2 Huttenlocker, A.K.; Sidor, C.A.; Smith, R.M.H. (2011). "A new specimen of Promoschorhynchus (Therapsida: Therocephalia: Akidnognathidae) from the Lower Triassic of South Africa and its implications for theriodont survivorship across the Permo-Triassic boundary". Journal of Vertebrate Paleontology 31 (2): 405–421. doi:10.1080/02724634.2011.546720.
- ↑ Sidor, C.A. (2001). "Simplification as a trend in synapsid cranial evolution". Evolution 55 (7): 1419–1442. doi:10.1554/0014-3820(2001)055[1419:saatis]2.0.co;2. PMID 11525465. External link in
|title=(help) - ↑ Huttenlocker, A. (2009). "An investigation into the cladistic relationships and monophyly of therocephalian therapsids (Amniota: Synapsida)". Zoological Journal of the Linnean Society 157 (4): 865–891. doi:10.1111/j.1096-3642.2009.00538.x.
- 1 2 Huttenlocker, A. K. (2014). "Body Size Reductions in Nonmammalian Eutheriodont Therapsids (Synapsida) during the End-Permian Mass Extinction". PLoS ONE 9 (2): e87553. doi:10.1371/journal.pone.0087553.
- ↑ Sigurdsen, T.; Huttenlocker, A. K.; Modesto, S. P.; Rowe, T. B.; Damiani, R. (2012). "Reassessment of the morphology and paleobiology of the therocephalian Tetracynodon darti (Therapsida), and the phylogenetic relationships of Baurioidea". Journal of Vertebrate Paleontology 32 (5): 1113. doi:10.1080/02724634.2012.688693.
Further reading
- Sigurdsen, T. (2006). "New features of the snout and orbit of a therocephalian therapsid from South Africa". Acta Palaeontologica Polonica 51 (1): 63–75.
- van den Heever, J.A. 1980. " On the validity of the therocephalian family Lycosuchidae (Reptilia, Therapsida)". Annals of the South African Museum 81: 111-125.
- van den Heever, J.A. 1994. "The cranial anatomy of the early Therocephalia (Amniota: Therapsida)." Annals of the University of Stellenbosch 1994: 1-59.
External links
| ||||||||||||||||||||||||||||||||||||||||||||||