Smilodon
| Smilodon Temporal range: Early Pleistocene to Early Holocene, 2.5–0.01 Ma | |
|---|---|
 | |
| S. fatalis skeleton at National Museum of Natural History, Washington, D.C. | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Carnivora |
| Family: | Felidae |
| Subfamily: | †Machairodontinae |
| Tribe: | †Smilodontini |
| Genus: | †Smilodon Lund, 1842 |
| Species | |
| Synonyms | |
|
Genus synonymy
Species synonymy
| |
Smilodon /ˈsmaɪlədɒn/, is an extinct genus of machairodont felid. It is perhaps one of the most famous prehistoric mammals, and the best known saber-toothed cat. Although commonly known as the saber-toothed tiger, it was not closely related to the tiger or other modern cats. Smilodon lived in the Americas during the Pleistocene epoch (2.5 mya–10,000 years ago). The genus was named in 1842, based on fossils from Brazil. Three species are recognized today: S. gracilis, S. fatalis and S. populator. The two latter species were probably descended from S. gracilis, which itself probably evolved from Megantereon. The largest collection of Smilodon fossils has been obtained from the La Brea Tar Pits.
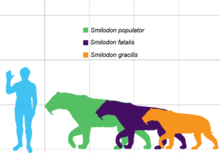
Overall, Smilodon was more robustly built than any extant cat, with particularly well-developed forelimbs and exceptionally long upper canines. Its jaw had a bigger gape than that of modern cats and its upper canines were slender and fragile, being adapted for precision killing. S. gracilis was the smallest species at 55 to 100 kg (120 to 220 lb) in weight. S. fatalis had a weight of 160 to 280 kg (350 to 620 lb) and height of 100 cm (39 in). Both of these species are mainly known from North America, but remains from South America have also been attributed to them. S. populator from South America is perhaps the largest known felid at 220 to 400 kg (490 to 880 lb) in weight and 120 cm (47 in) in height. The coat pattern of Smilodon is unknown, but it has been artistically restored with plain or spotted patterns.
In North America, Smilodon hunted large herbivores such as bison and camels and it remained successful even when encountering new prey species in South America. Smilodon is thought to have killed its prey by holding it still with its forelimbs and biting it, but it is unclear in what manner the bite itself was delivered. Scientists debate whether Smilodon had a social or a solitary lifestyle; analysis of modern predator behavior as well as of Smilodon's fossil remains could be construed to lend support to either view. Smilodon probably lived in closed habitats such as forests and bush, which would have provided cover for ambushing prey. Smilodon died out at the same time that most North and South American megafauna disappeared, about 10,000 years ago. Its reliance on large animals has been proposed as the cause of its extinction, along with climate change and competition with other species, but the exact cause is unknown.
Taxonomy

During the 1830s, Danish naturalist Peter Wilhelm Lund and his assistants collected fossils in the calcareous caves near the small town of Lagoa Santa, Minas Gerais, Brazil. Among the thousands of fossils found, he recognized a few isolated cheek teeth as belonging to a hyena, which he named Hyaena neogaea in 1839. After more material was found (including canine teeth and foot bones), Lund concluded the fossils instead belonged to a distinct genus of felid, though transitional to the hyenas. He stated it would have matched the largest modern predators in size, and was more robust than any modern cat. Lund originally wanted to name the new genus Hyænodon, but realizing this had recently become preoccupied by another prehistoric predator, he instead named it Smilodon populator in 1842. He explained the Ancient Greek meaning of Smilodon as σμίλη (smilē), a scalpel or two-edged knife, and ὀδoύς (odoús), tooth. This has also been translated as "tooth shaped like double-edged knife". He explained the species name populator as "the destroyer", which has also been translated as "he who brings devastation". By 1846, Lund had acquired nearly every part of the skeleton (from different individuals), and more specimens were found in neighboring countries by other collectors in the following years.[1][2] Though some later authors used Lund's original species name neogaea instead of populator, it is now considered an invalid nomen nudum ("naked name"), as it was not accompanied with a proper description and no type specimens were designated.[3] Some South American specimens have been referred to other genera, subgenera, species, and subspecies, such as Smilodontidion riggii, Smilodon (Prosmilodon) ensenadensis, and S. bonaeriensis, but these are now thought to be junior synonyms of S. populator.[4]

Fossils of Smilodon were discovered in North America from the second half of the 19th century onwards.[1] In 1869, American paleontologist Joseph Leidy described a maxilla fragment with a molar, which had been discovered in a petroleum bed in Hardin County, Texas. He referred the specimen to the genus Felis (which was then used for most cats, extant as well as extinct) but found it distinct enough to be part of its own subgenus, as F. (Trucifelis) fatalis.[5] The species name means "fate" or "destiny", but it is thought Leidy intended it to mean "fatal".[6] In an 1880 article about extinct American cats, American paleontologist Edward Drinker Cope pointed out that the F. fatalis molar was identical to that of Smilodon, and he proposed the new combination S. fatalis.[7] Most North American finds were scanty until excavations began in the La Brea Tar Pits in Los Angeles, where hundreds of individuals of S. fatalis have been found since 1875.[1] S. fatalis has junior synonyms such as S. mercerii, S. floridanus, and S. californicus.[4] American paleontologist Annalisa Berta considered the holotype of S. fatalis too incomplete to be an adequate type specimen, and the species has at times been proposed to be a junior synonym of S. populator.[3] Swedish paleontologists Björn Kurtén and Lars Werdelin supported the distinctness of the two species in 1990.[8]
In his 1880 article about extinct cats, Cope also named a third species of Smilodon, S. gracilis. The species was based on a partial canine, which had been obtained in a cave near the Schuylkill River in Pennsylvania. Cope found the canine to be distinct from that of the other Smilodon species due to its smaller size and more compressed base.[7] Its specific name refers to the species' lighter build.[9] This species is known from fewer and less complete remains than the other members of the genus.[10] S. gracilis has at times been considered part of genera such as Megantereon and Ischyrosmilus.[11] S. populator, S. fatalis and S. gracilis are currently considered the only valid species of Smilodon, and features used to define most of their junior synonyms have been dismissed as variation between individuals of the same species (intraspecific variation).[4][3] One of the most famous of prehistoric mammals, Smilodon has often been featured in popular media and is the state fossil of California.[1]
Evolution

Long the most completely known saber-toothed cat, Smilodon is still one of the best-known members of the group, to the point where the two concepts have been confused. The term "saber-tooth" refers to an ecomorph consisting of various groups of extinct predatory synapsids (mammals and close relatives), which convergently evolved extremely long maxillary canines, as well as adaptations to the skull and skeleton related to their use. This includes members of Gorgonopsia, Thylacosmilidae, Machaeroidinae, Nimravidae, Barbourofelidae, and Felidae.[1][12] Within the family Felidae (true cats), members of the subfamily Machairodontinae are referred to as saber-toothed cats, and this group is itself divided into three tribes: Metailurini (false saber-tooths), Homotherini (scimitar-toothed cats), and Smilodontini (dirk-toothed cats), to which Smilodon belongs.[4] Members of Smilodontini are defined by their long slender canines with fine to no serrations, whereas Homotherini are typified by shorter, broad, and more flattened canines, with coarser serrations.[13] Members of Metailurini were less specialized and had shorter, less flattened canines, and are not recognized as members of Machairodontinae by some researchers.[4]

The earliest felids are known from the Oligocene of Europe, such as Proailurus, and the earliest one with saber-tooth features is the Miocene genus Pseudaelurus.[4] The skull and mandible morphology of the earliest saber-toothed cats was similar to that of the modern clouded leopards (Neofelis). The lineage further adapted to the precision killing of large animals by developing elongated canine teeth and wider gapes, in the process sacrificing high bite force.[14] As their canines became longer, the bodies of the cats became more robust for immobilizing prey.[13] In derived smilodontins and homotherins, the lumbar region of the spine and the tail became shortened, as did the hind limbs.[4] The earliest species of Smilodon is S. gracilis, which existed from 2.5 million to 500,000 years ago (early Blancan to Irvingtonian ages) and was the successor in North America of Megantereon, from which it probably evolved. Megantereon itself had entered North America from Eurasia during the Pliocene, along with Homotherium. S. gracilis reached the northern regions of South America in the early Pleistocene as part of the Great American Interchange.[15][13] The younger Smilodon species are probably derived from S. gracilis.[16] S. fatalis existed 1.6 million–10,000 years ago (late Irvingtonian to Rancholabrean ages), and replaced S. gracilis in North America.[8] S. populator existed 1 million–10,000 years ago (Ensenadan to Lujanian ages); it occurred in the eastern parts of South America.[17]
Despite the colloquial name "saber-toothed tiger", Smilodon is not closely related to the modern tiger (which belongs in the subfamily Pantherinae), or any other extant felid.[18] An 1992 ancient DNA analysis suggested that Smilodon should be grouped with modern cats (subfamilies Felinae and Pantherinae).[19] However, a 2005 study found that Smilodon belonged to a separate lineage.[20] A study published in 2006 confirmed this, showing that the Machairodontinae diverged early from the ancestors of modern cats and were not closely related to any living species.[21] The following cladogram based on fossils and DNA analysis shows the placement of Smilodon among extinct and extant felids, after Rincón and colleagues, 2011:[15]

| Felidae |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |
Description
Smilodon was around the size of modern big cats, but was more robustly built. It had a reduced lumbar region, high scapula, short tail, and broad limbs with relatively short feet.[22][23] Smilodon is most famous for its relatively long canine teeth, which are the longest found in the saber-toothed cats, at about 28 cm (11 in) long in the largest species, S. populator.[22] The canines were slender and had fine serrations on the front and back side.[24] The skull was robustly proportioned and the muzzle was short and broad. The cheek bones (zygomata) were deep and widely arched, the sagittal crest was prominent, and the frontal region was slightly convex. The mandible had a flange on each side of the front. The upper incisors were large, sharp, and slanted forwards. There was a diastema (gap) between the incisors and molars of the mandible. The lower incisors were broad, recurved, and placed in a straight line across. The p3 premolar tooth of the mandible was present in most early specimens, but lost in later specimens; it was only present in 6% of the La Brea sample.[3] There is some dispute over whether Smilodon was sexually dimorphic. Some studies of S. fatalis fossils have found little difference between the sexes.[25][26] Conversely, a 2012 study found that, while fossils of S. fatalis show less variation in size among individuals than modern Panthera, they do appear to show the same difference between the sexes in some traits.[27]

S. gracilis was the smallest species, estimated at 55 to 100 kg (120 to 220 lb) in weight, about the size of a jaguar. It was similar to its predecessor Megantereon of the same size, but its dentition and skull were more advanced, approaching S. fatalis.[28][4] S. fatalis was intermediate in size between S. gracilis and S. populator.[22] It ranged from 160 to 280 kg (350 to 620 lb).[28] and reached a shoulder height of 100 cm (39 in) and body length of 175 cm (69 in).[29] It was similar to a lion in dimensions, but was more robust and muscular, and therefore had a larger body mass. Its skull was also similar to that of Megantereon, though more massive and with larger canines.[4] S. populator was perhaps the largest known felid, with a body mass range of 220 to 400 kg (490 to 880 lb),[28] and one estimate suggesting up to 470 kg (1,040 lb).<ref name = ""Sorkin">Sorkin, B. (2008). "A biomechanical constraint on body mass in terrestrial mammalian predators". Lethaia 41 (4): 333–347. doi:10.1111/j.1502-3931.2007.00091.x.</ref> It stood at a shoulder height of 120 cm (47 in).[22] Compared to S. fatalis, S. populator was more robust and had a more elongated and narrow skull with a straighter upper profile, higher positioned nasal bones, a more vertical occiput, more massive metapodials and slightly longer forelimbs relative to hindlimbs.[4][8]

Traditionally, saber-toothed cats have been artistically restored with external features similar to those of extant felids, by artists such as Charles R. Knight in collaboration with various paleontologists in the early 20th century.[30] In 1969, paleontologist G. J. Miller instead proposed that Smilodon would have looked very different from a typical cat and similar to a bulldog, with a lower lip line (to allow its mouth to open wide without tearing the facial tissues), a more retracted nose and lower-placed ears.[31] Paleoartist Mauricio Antón and coauthors disputed this in 1998 and maintained that the facial features of Smilodon were overall not very different from those of other cats. Antón noted that modern animals like the hippopotamus are able to achieve a wide gap without tearing tissue by the moderate folding of the orbicularis oris muscle, and such a muscle configuration exists in modern large felids.[32] Antón stated that extant phylogenetic bracketing (where the features of the closest extant relatives of a fossil taxon are used as reference) is the most reliable way of restoring the life-appearance of prehistoric animals, and the cat-like Smilodon restorations by Knight are therefore still accurate.[30]
Smilodon and other saber-toothed cats have been reconstructed with both plain-colored coats and with spotted patterns (which appears to be the ancestral condition for feliforms), both of which are considered possible.[30] Studies of modern cat species have found that species that live in the open tend to have uniform coats while those that live in more vegetated habitats have more markings. However, exceptions to both cases exist.[33] Some coat features, such as the manes of male lions or the stripes of the tiger, are too unusual to predict from fossils.[30]
Paleobiology
Predatory behavior

An apex predator, Smilodon primarily hunted large mammals. Isotopes preserved in the bones of S. fatalis in the La Brea Tar Pits reveal that ruminants like bison (Bison antiquus, which was much larger than the modern American bison) and camels (Camelops) were most commonly taken by the cats there.[34] In addition, isotopes preserved in the tooth enamel of S. gracilis specimens from Florida show that this species fed on the pig-like Platygonus and the llama-like Hemiauchenia.[35] Isotopic studies of dire wolf (Canis dirus) and American lion (Panthera leo atrox) bones show an overlap with S. fatalis in prey, which suggests that they were competitors.[34] The availability of prey in the Rancho La Brea area was likely comparable to modern East Africa.[36] As Smilodon migrated to South America, its diet changed; bison were absent, the horses and proboscideans were different, and native ungulates such as toxodonts and litopterns were completely unfamiliar, yet S. populator thrived as well there as its relatives in North America.[13] The differences between the North and South American species may be due to the difference in prey between the two continents.[8] Smilodon probably avoided eating bone and would have left enough food for scavengers.[37] Smilodon itself may have scavenged dire wolf kills.[38] It has been suggested that Smilodon was a scavenger that mainly used its canines for display to assert dominance over carcasses, but this theory is not supported today and no modern terrestrial mammals are pure scavengers.[39]
The brain of Smilodon had sulcal patterns similar to modern cats, which suggests an increased complexity of the regions that control the sense of hearing, sight, and coordination of the limbs. Felid saber-tooths in general had relatively small eyes that were not as forward-facing as those of modern cats, which have good binocular vision to help them move in trees.[39] Smilodon was likely an ambush predator that concealed itself in dense vegetation.[40] The heel bone of Smilodon was fairly long, which suggests it was a good jumper.[22] Its well-developed flexor and extensor muscles in its forearms probably enabled it to pull down, and securely hold down, large prey. Analysis of the cross-sections of S. fatalis humeri indicated that they were strengthened by cortical thickening to such an extent that they would have been able to sustain greater loading than those of extant big cats, or of the extinct American lion. However, the thickening of S. fatalis femurs was within the range of extant felids.[41] As its canines were fragile and could not have bitten into bone, these cats did not use their long teeth while taking down prey, due to the risk of breaking, and had to subdue and restrain their prey so they could use the teeth. By contrast, modern cats can subdue and kill large prey with a slow suffocating bite.[41]

Debate continues as to how Smilodon killed its prey. Traditionally, the most popular theory is that the cat delivered a deep stabbing bite or open-jawed stabbing thrust to the throat, generally cutting through the jugular vein and/or the trachea and thus killing the prey very quickly.[41][42] Alternatively, it may have used its canines to puncture the thoracic wall of its prey (a larger, easier target) with a closed-mouth stab, creating a condition of pneumothorax (collapsed lungs).[43] Another hypothesis suggests that Smilodon targeted the belly of its prey. This is disputed, however, as the curvature of their prey's belly would likely have prevented the cat from getting a good bite or stab.[44] Whether Smilodon generally used its canines to deliver a point-to-point bite, open-jawed stab or closed-jawed stab is unclear.[43] In regard to how Smilodon delivered its bite, the "canine shear-bite" hypothesis has been favored, where flexion of the neck and rotation of the skull assisted in biting the prey, but this may be mechanically impossible. The mandibular flanges may have helped resist bending forces when the mandible was pulled against the hide of a prey.[45] The protruding incisors were arranged in an arch, and were used to hold the prey still and stabilize it while the canine bite was delivered. The contact surface between the canine crown and the gum was enlarged, which helped stabilize the tooth and helped the cat sense when the tooth had penetrated to its maximum extent. Since saber-toothed cats generally had a relatively large infraorbital foramen (opening) in the skull, which housed nerves associated with the whiskers, it has been suggested the improved senses would have helped the cats' precision when biting outside their field of vision, and thereby prevent breakage of the canines. The blade-like carnassial teeth were used to cut skin to access the meat, and the reduced molars suggest that they were less adapted for crushing bones than modern cats.[39] As the food of modern cats enters the mouth through the side while cutting with the carnassials, not the front between the canines, the animals do not need to gape widely, so the canines of Smilodon would likewise not have been a hindrance when feeding.[32]

Despite being more powerfully built than other large cats, Smilodon had a weaker bite. Modern big cats have more pronounced zygomatic arches, while these were smaller in Smilodon, which restricted the thickness and therefore power of the temporalis muscles and thus reduced Smilodon's bite force. Analysis of its narrow jaws indicates that it could produce a bite only a third as strong as that of a lion.[46] There seems to a be a general rule that the saber-toothed cats with the largest canines had proportionally weaker bites. However, analyses of canine bending strength (the ability of the canine teeth to resist bending forces without breaking) and bite forces indicate that the saber-toothed cats' teeth were stronger relative to the bite force than those of modern big cats.[47] In addition, Smilodon's gape could have reached almost 120 degrees,[48] while that of the modern lion reaches 65 degrees.[49] This made the gape wide enough to allow Smilodon to grasp large prey despite the long canines.[32]
Natural traps
Many Smilodon specimens have been excavated from asphalt seeps that once acted as natural carnivore traps. Animals were accidentally trapped in the seeps and became bait for predators that came to scavenge, but these were then trapped themselves. The best-known of such traps are the La Brea Tar Pits of Los Angeles, which have produced the largest sample of saber-toothed cat fossils in the world. The sediments of the pits there were accumulated 40,000 to 10,000 years ago, in the Late Pleistocene. Though the trapped animals were buried quickly, predators often managed to remove limb bones from them, but they were themselves often trapped and then scavenged by other predators; 90% of the excavated bones belonged to predators.[50]
The Talara Tar Seeps in Peru represent a similar scenario, and have also produced fossils of Smilodon. Unlike in La Brea, many of the bones were broken or show signs of weathering. This may have been because the layers were shallower, so the thrashing of trapped animals damaged the bones of previously trapped animals. Many of the carnivores at Talara were juveniles, possibly indicating that inexperienced and less fit animals had a greater chance of being trapped. Though Lund thought accumulations of Smilodon and herbivore fossils in the Lagoa Santa Caves were due to the cats using the caves as dens, these are probably the result of animals dying on the surface, and water currents subsequently dragging their bones to the floor of the cave, but some individuals may also have died after becoming lost in the caves.[50]
Social life

Scientists debate whether Smilodon was social. One study of African predators found that social predators like lions and spotted hyenas respond more to the distress calls of prey than solitary species. Since S. fatalis fossils are common at the La Brea Tar Pits, and were likely attracted by the distress calls of stuck prey, this could mean that this species was social as well.[51] One critical study claims that the study neglects other factors, such as body mass (heavier animals are more likely to get stuck than lighter ones), intelligence (some social animals, like the American lion, may have avoided the tar because they were better able to recognize the hazard), lack of visual and olfactory lures, the type of audio lure, and the length of the distress calls (the actual distress calls of the trapped prey animals would have lasted longer than the calls used in the study). The author of that study ponders what predators would have responded if the recordings were played in India, where the otherwise solitary tigers are known to aggregate around a single carcass.[52] The authors of the original study responded that though effects of the calls in the tar pits and the playback experiments would not be identical, this would not be enough to overturn their conclusions. In addition, they stated that weight and intelligence would not likely affect the results as lighter carnivores are far more numerous than heavy herbivores and the social (and seemingly intelligent) dire wolf is also found in the pits.[53]

Another argument for sociality is based on the healed injuries in several Smilodon fossils, which would suggest that the animals needed others to provide it food.[54] This argument has been questioned, as cats can recover quickly from even severe bone damage and an injured Smilodon could survive if it had access to water.[55] The brain of Smilodon was relatively small compared to other cat species. Some researchers have argued that Smilodon's brain would have been too small for it to have been a social animal.[56] However, an analysis of brain size in living big cats found no correlation between brain size and sociality.[57] Another argument against Smilodon being social is that being an ambush hunter in closed habitat would likely have made group-living unnecessary, as in most modern cats.[55] Yet it has also been proposed that being the largest predator in an environment comparable to the savanna of Africa, Smilodon may have had a social structure similar to modern lions, which possibly live in groups primarily to defend optimal territory from other lions (lions are the only social big cats today).[39]
Whether Smilodon was sexually dimorphic has implications for its reproductive behavior. Based on their conclusions that Smilodon fatalis had no sexual dimorphism, Van Valenburgh and Sacco (2002) suggested that, if the cats were social, they would likely have lived in monogamous pairs (along with offspring) with no intense competition among males for females.[25] Likewise, Meachen-Samuels and Binder (2010) concluded that aggression between males was less pronounced in S. fatalis than in the American lion.[26] However, Christiansen and Harris (2012) found that, as S. fatalis did exhibit some sexual dimorphism, there would have been evolutionary selection for competition between males.[27] The structure of the hyoid bones suggest that Smilodon communicated by roaring, like modern big cats.[58]
Development

Smilodon started developing its adult saber-teeth when the animal turned one-and-a-half years of age, shortly after the completion of the eruption of the cat's baby teeth. Both baby and adult canines would be present side by side in the mouth for an 11-month period, and the muscles used in making the powerful bite were developed at about one-and-a-half years old as well, eight months earlier than in a modern lion. After Smilodon turned 20 months of age, the infant teeth were shed while the adult canines grew in, a process that continued until the cats reached 3 years of age, later than for modern species of big cat. Juvenile and adolescent Smilodon specimens are extremely rare at Rancho La Brea, where the study was performed, indicating that they remained hidden or at denning sites during hunts, and depended on parental care while their canines were developing.[59][60] The teeth of S. fatalis reached their full size in 18 months at a growth rate of 7 mm (0.3 in) per month.[61]
Paleopathology
Several Smilodon fossils show signs of ankylosing spondylitis, hyperostosis and trauma;[62] some also had arthritis, which gave them fused vertebrae. One study of 1,000 Smilodon skulls found that 30% of them had eroded parietal bones, which is where the largest jaw muscles attach. They also showed signs of microfractures, and the weakening and thinning of bones possibly caused by mechanical stress from the constant need to make stabbing motions with the canines.[63] Bony growths where the deltoid muscle inserted in the humerus is a common pathology for a La Brea specimen, which was probably due to repeated strain when Smilodon attempted to pull down prey with its forelimbs. Sternum injuries are also common, probably due to collision with prey. Some bones also show evidence of having been bitten by other Smilodon, with one skull showing an unhealed wound from a canine, seemingly fatal, and a scapula from the same site that had been pierced by a canine.[39]
Distribution and habitat

Smilodon lived during the Pleistocene epoch (2.5 mya–10,000 years ago), and was perhaps the most recent of the saber-toothed cats.[22] Smilodon probably lived in a closed habitat such as forest or bush.[64] Fossils of the genus have been found throughout the Americas.[3] In North America, the varied habitat supported other saber-toothed cats in addition to Smilodon, such as Homotherium and Xenosmilus; the habitat here varied from subtropical forests and savannah in the south, to treeless mammoth steppes in the north. Smilodon inhabited the temperate latitudes of North America, where the mosaic vegetation of woods, shrubs, and grasses in the southwest supported large herbivores such as horses, bison, antelope, deer, camels, mammoths, mastodons, and ground sloths. Other large carnivores include dire wolves and the American lion.[13][50]
Smilodon gracilis entered South America during the early to middle Pleistocene, where it probably gave rise to S. populator, which lived in the eastern part of the continent. S. fatalis also entered western South America in the late Pleistocene.[8][15] The American interchange resulted in a mix of native and invasive species sharing the prairies and woodlands in South America; North American herbivores included proboscideans, horses, camelids and deer, South American herbivores included toxodonts, litopterns, ground sloths, and glyptodonts. Native metatherian predators (including the saber-toothed thylacosmilids) had gone extinct by the Pliocene, and were replaced by North American carnivores such as canids, bears, and large cats. S. populator was very successful here, while Homotherium never became widespread. The extinction of the thylacosmilids has been attributed to competition with Smilodon, but this is probably incorrect, as they seem to have disappeared before the arrival of the large cats. The phorusrhacid "terror birds" may have dominated the large predator niche in South America until Smilodon arrived.[13]
Extinction

Along with most of the Pleistocene megafauna, Smilodon went extinct 10,000 years ago in the Quaternary extinction event. Its extinction has been linked to the decline and extinction of large herbivores, which were replaced by smaller and more agile ones like deer. Hence, Smilodon could have been too specialized at hunting large prey and may have been unable to adapt.[41] However, a 2012 study of Smilodon tooth wear found no evidence that they were limited by food resources.[65] Other explanations include climate change and competition with humans (which entered the Americas around the time Smilodon disappeared), or a combination of several factors, all of which apply to the general Pleistocene extinction event, rather than specifically to the extinction of the saber-toothed cats.[66]
Some early writers theorized that the last saber-toothed cats, Smilodon and Homotherium, went extinct through competition with the faster and more generalized felids that replaced them. It was even proposed that the saber-toothed predators were inferior to modern cats, as the ever-growing canines were thought to inhibit their owners from feeding properly. Yet fast felids such as the American lion and the American cheetah also went extinct during the Late Pleistocene. The fact that saber-teeth evolved many times in unrelated lineages also attests to the success of this feature.[66]
References
- 1 2 3 4 5 Antón 2013, pp. 3–26.
- ↑ Lund, P. W. (1842). Blik paa Brasiliens Dyreverden för sidste Jordomvæltning (in Danish). Copenhagen: Det Kongelige Danske Videnskabernes Selskabs Naturvidenskabelige of Matematiske Afhandlinger. pp. 54–57.
- 1 2 3 4 5 Berta, A. (1985). "The status of Smilodon in North and South America" (PDF). Contributions in Science, Natural History Museum of Los Angeles County 370: 1–15.
- 1 2 3 4 5 6 7 8 9 10 Antón 2013, pp. 108–154.
- ↑ Leidy, J. (1869). "The extinct mammalian fauna of Dakota and Nebraska: Including an account of some allied forms from other localities, together with a synopsis of the mammalian remains of North America". Journal of the Academy of Natural Sciences of Philadelphia 7: 366–367. doi:10.5962/bhl.title.20910.
- ↑ Hulbert, R. C. (2013-04-23). "Smilodon fatalis Leidy, 1868". Florida Museum of Natural History. Retrieved 2013-05-01.
- 1 2 Cope, E. D. (December 1880). "On the extinct cats of America". The American Naturalist 14 (12): 833–858. doi:10.1086/272672. JSTOR 2449549.
- 1 2 3 4 5 Kurtén, B.; Werdelin, L. (1990). "Relationships between North and South American Smilodon". Journal of Vertebrate Paleontology 10 (2): 158–169. doi:10.1080/02724634.1990.10011804. JSTOR 4523312.
- ↑ Kurten, B.; Anderson, E. (1980). Pleistocene Mammals of North America. New York: Columbia University Press. pp. 186–188. ISBN 978-0-231-03733-4.
- ↑ Berta, A. (1987). "The sabercat Smilodon gracilis from Florida and a discussion of its relationships (Mammalia, Felidae, Smilodontini)". Bulletin of the Florida State Museum 31: 1–63.
- ↑ Churcher, C. S. (1984). "The status of Smilodontopsis (Brown, 1908) and Ischyrosmilus (Merriam, 1918): a taxonomic review of two genera of sabretooth cats (Felidae, Machairodontinae)". Royal Ontario Museum Life Sciences Contributions 140: 14–34. doi:10.5962/bhl.title.52222. ISBN 978-0-88854-305-9.
- ↑ Meehan, T. J.; Martin, L. D. (2003). "Extinction and re-evolution of similar adaptive types (ecomorphs) in Cenozoic North American ungulates and carnivores reflect van der Hammen's cycles". Die Naturwissenschaften 90 (3): 131–135. Bibcode:2003NW.....90..131M. doi:10.1007/s00114-002-0392-1 (inactive 2016-01-02). PMID 12649755.
- 1 2 3 4 5 6 Antón 2013, pp. 65–76.
- ↑ Christiansen, P. (2008). "Evolution of skull and mandible shape in cats (Carnivora: Felidae)". PLoS ONE 3 (7): e2807. Bibcode:2008PLoSO...3.2807C. doi:10.1371/journal.pone.0002807.
- 1 2 3 Rincón, A.; Prevosti, F.; Parra, G. (2011). "New saber-toothed cat records (Felidae: Machairodontinae) for the Pleistocene of Venezuela, and the Great American Biotic Interchange". Journal of Vertebrate Paleontology 31 (2): 468–478. doi:10.1080/02724634.2011.550366. JSTOR 25835839.
- ↑ Ascanio, D.; Rincón R. (2006). "A first record of the Pleistocene saber-toothed cat Smilodon populator Lund, 1842 (Carnivora: Felidae: Machairodontinae) from Venezuela". Asociación Paleontologica Argentina 43 (2): 499–501. ISSN 1851-8044.
- ↑ Mariela Cordeiro de Castro, Max Cardoso Langer (2008). "New postcranial remains of Smilodon populator Lund, 1842 from South-Central Brazil" (PDF). Revista Brasileira de Paleontologia 11 (3): 199–206. doi:10.4072/rbp.2008.3.06.
- ↑ "What Is a Sabertooth?". University of California Museum of Paleontology. December 2005. Retrieved 2012-06-12.
- ↑ Janczewski, D. N.; Yuhki, N.; Gilbert, D. A.; Jefferson, G. T.; O'Brien, S. J. (1992). "Molecular phylogenetic inference from saber-toothed cat fossils of Rancho La Brea". Proceedings of the National Academy of Sciences 89 (20): 9769–9773. Bibcode:1992PNAS...89.9769J. doi:10.1073/pnas.89.20.9769.
- ↑ Barnett, R; Barnes, I.; Phillips, M. J.; Martin, L. D.; Harington, C. R.; Leonard, J. A.; Cooper, A. (2005). "Evolution of the extinct sabretooths and the American cheetah-like cat". Current Biology 15 (15): R589–R590. doi:10.1016/j.cub.2005.07.052. PMID 16085477.
- ↑ van den Hoek Ostende, L. W.; Morlo, M.; Nagel, D. (2006). "Majestic killers: the sabre-toothed cats (Fossils explained 52)". Geology Today 22 (4): 150–157. doi:10.1111/j.1365-2451.2006.00572.x.
- 1 2 3 4 5 6 Turner, A.; Antón, M. (1997). The Big Cats and Their Fossil Relatives: An Illustrated Guide to Their Evolution and Natural History. Columbia University Press. pp. 57–58, 67–68. ISBN 978-0-231-10229-2. OCLC 34283113.
- ↑ "What Is a Sabertooth?". University of California Museum of Paleontology. Retrieved 2013-04-08.
- ↑ Slater, G. J.; Valkenburgh, B. V. (2008). "Long in the tooth: evolution of sabertooth cat cranial shape". Paleobiology 34 (3): 403–419. doi:10.1666/07061.1. ISSN 0094-8373.
- 1 2 Van Valkenburgh, B.; Sacco, T. (2002). "Sexual dimorphism, social behavior and intrasexual competition in large Pleistocene carnivorans". Journal of Vertebrate Paleontology 22 (1): 164–169. doi:10.1671/0272-4634(2002)022[0164:sdsbai]2.0.co;2. JSTOR 4524203.
- 1 2 Meachen-Samuels, J.; Binder, W. (2010). "Sexual dimorphism and ontogenetic growth in the American lion and sabertoothed cat from Rancho La Brea". Journal of Zoology 280 (3): 271–279. doi:10.1111/j.1469-7998.2009.00659.x.
- 1 2 Christiansen, P.; Harris J. M. (2012). "Variation in craniomandibular morphology and sexual dimorphism in Pantherines and the sabercat Smilodon fatalis". PLoS ONE 7 (10): e48352. Bibcode:2012PLoSO...748352C. doi:10.1371/journal.pone.0048352.
- 1 2 3 Christiansen, P.; Harris, J. M. (2005). "Body size of Smilodon (Mammalia: Felidae)". Journal of Morphology 266 (3): 369–384. doi:10.1002/jmor.10384. PMID 16235255.
- ↑ "Saber-Toothed Cat, Smilodon fatalis". San Diego Zoo Global. January 2009. Retrieved 2013-05-07.
- 1 2 3 4 Antón 2013, pp. 157–176.
- ↑ Miller, G. J. (1969). "A new hypothesis to explain the method of food ingestion used by Smilodon californicus Bovard". Tebiwa 12: 9–19.
- 1 2 3 Antón, M.; García-Perea, R.; Turner, A. (1998). "Reconstructed facial appearance of the sabretoothed felid Smilodon". Zoological Journal of the Linnean Society 124 (4): 369–386. doi:10.1111/j.1096-3642.1998.tb00582.x.
- ↑ Allen, W. L.; Cuthill, I. C.; Scott-Samuel, N. E.; Baddeley, R. (2010). "Why the leopard got its spots: relating pattern development to ecology in felids". Proceedings of the Royal Society B 278 (1710): 1373–1380. doi:10.1098/rspb.2010.1734. PMC 3061134. PMID 20961899.
- 1 2 Coltrain, J. B.; Harris, J. M.; Cerling, T. E.; Ehleringer, J. R.; Dearing, M-D.; Ward, J.; Allen, J. (2004). "Rancho La Brea stable isotope biogeochemistry and its implications for the palaeoecology of late Pleistocene, coastal southern California". Palaeogeography, Palaeoclimatology, Palaeoecology 205 (3–4): 199–219. doi:10.1016/j.palaeo.2003.12.008.
- ↑ Fennec, R. S. (2005). "Growth rate and duration of growth in the adult canine of S. gracilis and inferences on diet through stable isotope analysis". Feranec Bull FLMNH 45 (4): 369–377.
- ↑ Vanvalkenburgh, B.; Hertel, F. (1993). "Tough times at la brea: tooth breakage in large carnivores of the late Pleistocene". Science 261 (5120): 456–459. Bibcode:1993Sci...261..456V. doi:10.1126/science.261.5120.456. PMID 17770024.
- ↑ Van Valkenburgh, B.; Teaford, M. F.; Walker, A. (1990). "Molar microwear and diet in large carnivores: inferences concerning diet in the sabretooth cat, Smilodon fatalis". Journal of Zoology 222 (2): 319–340. doi:10.1111/j.1469-7998.1990.tb05680.x.
- ↑ Van Valkenburgh, B. (1991). "Iterative evolution of hypercarnivory in canids (Mammalia: Carnivora): evolutionary interactions among sympatric predators". Paleobiology 17 (4): 340–362. JSTOR 2400749.
- 1 2 3 4 5 Antón 2013, pp. 176–216.
- ↑ Gonyea, W. J. (1976). "Behavioral implications of saber-toothed felid morphology". Paleobiology 2 (4): 332–342. JSTOR 2400172.
- 1 2 3 4 Meachen-Samuels, J. A.; Van Valkenburgh, B. (2010). "Radiographs reveal exceptional forelimb strength in the sabertooth cat, Smilodon fatalis". PLoS ONE 5 (7): e11412. doi:10.1371/journal.pone.0011412. ISSN 1932-6203. PMID 20625398.
- ↑ McHenry, C. R.; Wroe, S.; Clausen, P. D.; Moreno, K.; Cunningham, E. (2007). "Supermodeled sabercat, predatory behavior in Smilodon fatalis revealed by high-resolution 3D computer simulation". PNAS 104 (41): 16010–16015. Bibcode:2007PNAS..10416010M. doi:10.1073/pnas.0706086104. PMC 2042153. PMID 17911253.
- 1 2 Wilson, T.; Wilson, D. E.; Zimanske, J. M. (2013). "Pneumothorax as a predatory goal for the sabertooth cat (Smilodon fatalis)". Open Journal of Animal Sciences 3 (1): 42–45. doi:10.4236/ojas.2013.31006. ISSN 2161-7597.
- ↑ Anyonge, W. (1996). "Microwear on canines and killing behavior in large carnivores: saber function in Smilodon fatalis" (PDF). Journal of Mammalogy 77 (4): 1059–1067. doi:10.2307/1382786. JSTOR 1382786.
- ↑ Macchiarelli, R.; Brown, J. G. (2014). "Jaw function in Smilodon fatalis: a reevaluation of the canine shear-bite and a proposal for a new forelimb-powered class 1 lever model". PLoS ONE 9 (10): e107456. doi:10.1371/journal.pone.0107456. ISSN 1932-6203. PMC 4182664. PMID 25272032.
- ↑ Hecht, J. (1 October 2007). "Sabre-tooth cat had a surprisingly delicate bite". New Scientist. The study used finite element analysis, a computerized technique common in engineering.
- ↑ Christiansen, P. (2007). "Comparative bite forces and canine bending strength in feline and sabretooth felids: implications for predatory ecology". Zoological Journal of the Linnean Society 151 (2): 423–437. doi:10.1111/j.1096-3642.2007.00321.x.
- ↑ Andersson, K.; Norman, D.; Werdelin, L.; (2011). "Sabretoothed carnivores and the killing of large prey". PLoS ONE 6 (10): e24971. Bibcode:2011PLoSO...624971A. doi:10.1371/journal.pone.0024971. PMC 3198467. PMID 22039403.
- ↑ Martin, L. D. (1980). "Functional morphology and the evolution of cats". Transactions of the Nebraska Academy of Sciences 8: 141–154.
- 1 2 3 Antón 2013, pp. 30–33.
- ↑ Carbone, C.; Maddox, T.; Funston, P. J.; Mills, M. G. L.; Grether, G. F.; Van Valkenburgh, B. (2009). "Parallels between playbacks and Pleistocene tar seeps suggest sociality in an extinct sabretooth cat, Smilodon". Biological Letters 5 (1): 81–85. doi:10.1098/rsbl.2008.0526. PMC 2657756. PMID 18957359.
- ↑ Kiffner, C. (2009). "Coincidence or evidence: was the sabretooth cat Smilodon social?". Biology Letters 5 (4): 561–562. doi:10.1098/rsbl.2009.0008. PMC 2781900. PMID 19443504.
- ↑ Van Valkenburgh, B.; Maddox, T.; Funston, P. J.; Mills, M. G. L.; Grether, G. F.; Carbone, C. (2009). "Sociality in Rancho La Brea Smilodon: arguments favour 'evidence' over 'coincidence'". Biology Letters 5 (4): 563–564. doi:10.1098/rsbl.2009.0261.
- ↑ Heald, F. (1989). "Injuries and diseases in Smilodon californicus". Journal of Vertebrate Paleontology (Supplement) 9: 24A.
- 1 2 McCall, S.; Naples, V.; Martin, L.; (2003). "Assessing behavior in extinct animals: was Smilodon social?". Brain, Behavior and Evolution 61 (3): 159–164. doi:10.1159/000069752. PMID 12697957.
- ↑ Radinsky, L. B. (1975). "Evolution of the felid brain". Brain, Behavior and Evolution 11 (3–4): 214–254. doi:10.1159/000123636. PMID 1181005.
- ↑ Yamaguchi, N.; Kitchener, A. C.; Gilissen, E.; MacDonald, D. W. (2009). "Brain size of the lion (Panthera leo) and the tiger (P. tigris): implications for intrageneric phylogeny, intraspecific differences and the effects of captivity". Biological Journal of the Linnean Society 98 (1): 85–93. doi:10.1111/j.1095-8312.2009.01249.x.
- ↑ Hearld, F.; Shaw, C. (1991). "Sabertooth Cats". In Seidensticker, J.; Lumpkin, S. The Great Cats. Rodale Pr. p. 26. ISBN 1-875137-90-4.
- ↑ "Dagger-like canines of saber-toothed cats took years to grow". ScienceDaily. 2015-07-01. Archived from the original on 2015-07-02. Retrieved 2015-07-02.
- ↑ Mihlbachler, M. C.; Wysocki, M. A.; Feranec, R. S.; Tseng, Z. J.; Bjornsson, C. S. (2015-07-01). "Using a novel absolute ontogenetic age determination technique to calculate the timing of tooth eruption in the saber-toothed cat, Smilodon fatalis". PLoS ONE 10 (7): e0129847. doi:10.1371/journal.pone.0129847. PMID 26132165.
- ↑ Feranec, R. C. (2004). "Isotopic evidence of saber-tooth development, growth rate, and diet from the adult canine of Smilodon fatalis from Rancho La Brea". Palaeogeography, Palaeoclimatology, Palaeoecology 206 (3–4): 303–310. doi:10.1016/j.palaeo.2004.01.009.
- ↑ Bjorkengren, A. G.; Sartoris, D. J.; Shermis, S.; Resnick, D. (1987). "Patterns of paravertebral ossification in the prehistoric saber-toothed cat". American Journal of Roentgenology 148 (4): 779–782. doi:10.2214/ajr.148.4.779. PMID 3103404.
- ↑ Duckler, G. L. (1997). "Parietal depressions in skulls of the extinct saber-toothed felid Smilodon fatalis: evidence of mechanical strain". Journal of Vertebrate Paleontology 17 (3): 600–609. doi:10.1080/02724634.1997.10011006.
- ↑ Meloro, C.; Elton, S.; Louys, J.; Bishop, L. C.; Ditchfield, P. (2013). "Cats in the forest: predicting habitat adaptations from humerus morphometry in extant and fossil Felidae (Carnivora)". Paleobiology 39 (3): 323–344. doi:10.1666/12001.
- ↑ DeSantis, L. R. G.; Schubert, B. W.; Scott, J. R.; Ungar, P. S. (2012). "Implications of diet for the extinction of saber-toothed cats and American lions". PLoS ONE 7 (12): e52453. Bibcode:2012PLoSO...752453D. doi:10.1371/journal.pone.0052453.
- 1 2 Antón 2013, pp. 217–230.
Bibliography
| Wikimedia Commons has media related to Smilodon. |
- Antón, M. (2013). Sabertooth (1 ed.). Bloomington: Indiana University Press. ISBN 978-0-253-01042-1.