Sarcocystis
| Sarcocystis | |
|---|---|
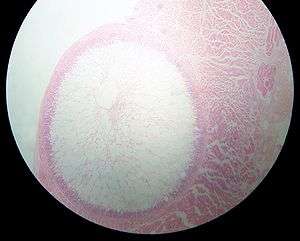 | |
| Sarcocystis cyst in a sheep oesophagus. The cyst is approximately 4 mm across. | |
| Scientific classification | |
| Domain: | Eukaryota |
| (unranked): | Sar |
| Superphylum: | Alveolata |
| Phylum: | Apicomplexa |
| Class: | Conoidasida |
| Subclass: | Coccidiasina |
| Order: | Eucoccidiorida |
| Suborder: | Eimeriorina |
| Family: | Sarcocystidae |
| Subfamily: | Sarcocystinae |
| Genus: | Sarcocystis Lankester, 1882 |
| Species | |
|
Sarcocystis accipitris | |
Sarcocystis is a genus of protozoa. Species in this genus are parasites, the majority infecting mammals, and some infecting reptiles and birds.
The life-cycle of a typical member of this genus involves two host species, a definitive host and an intermediate host. Often the definitive host is a predator and the intermediate host is its prey. The parasite reproduces sexually in the gut of the definitive host, is passed with the feces and ingested by the intermediate host. There it eventually enters muscle tissue. When the intermediate host is eaten by the definitive host, the cycle is completed. The definitive host usually does not show any symptoms of infection, but the intermediate host does.
There are about 130 recognized species in this genus. Revision of the taxonomy of the genus is ongoing, and it is possible that all the currently recognised species may in fact be a much smaller number of species that can infect multiple hosts.
The name Sarcocystis is dervived from Greek: sarx = flesh and kystis = bladder.
History
The organism was first recognised in a mouse by Miescher in 1843.[1] His findings were not recognised as a protist initially and the literature referred to the structures he described as "Miescher's Tubules". Incidentally Miescher's son — Johann Friedrich Miescher — discovered DNA. Similar structures were found in pig muscle in 1865 but these remained unnamed until 1899 when the name Sarcocystis meischeriana was proposed to identify them.
Initially it was unclear whether these organisms were fungi or protozoa. This uncertainty was resolved in 1967 when electron microscopic studies showed that these organism were protozoa, related to Toxoplasma and Eimeria. The life cycle remained unknown until 1970 when bradyzoites from sarcocysts in bird muscles were inoculated into cultured mammalian cells and underwent development into sexual stages and oocysts. Transmission studies with the three morphotypes found in cattle then considered a single species - Sarcoplasma fusiformis - in dogs, cats and humans revealed that these were three different species now named Sarcoplasma bovicanis, Sarcoplasma bovifelis and Sarcoplasma bovihominis.
Life cycle
The heteroxenous (more than one obligatory host in its life cycle) life cycle of these apicomplexan parasites remained obscure until 1972 when the prey-predator relationship of its definitive and intermediate hosts was recognised. The life cycles of about 60 of these species is now known.
In outline gametogony and sporogony occur in the intestine of the definitive host while both schizogony which occurs in various tissues and the formation of sarcocysts (containing bradyzoites and metrocytes) occurs principally in the muscles of the intermediate host. In some cases a single species may act as both the definitive and intermediate host.
Oocysts are passed in the feces of an infected definitive host. The oocyst undergoes sporogony creating two sporocysts. Once this is complete the oocyst itself undergoes lysis releasing the sporocysts into the environment. Sporocysts typically contain 4 sporozoites and measure 15-19 by 8-10 micrometres.
An intermediate host such as a cow or pig then ingests a sporocyst. Sporozoites are then released in the body and migrate to vessels where they undergo the first two generation of asexual reproduction. These rounds result in the development of meronts. This stage lasts about 15 to 16 days after ingestion of sporocysts. Merozoites emerge from the second generation meronts and enter the mononucleate cells where they develop by endodyogeny. Subsequent generations of merozoites develop downstream in the direction of blood flow to arterioles, capillaries, venules, and veins throughout the body subsequently developing into the final asexual generation in muscles.
Merozoites entering muscle cells round up to form metrocytes and initiate sarcocyst formation. Sarcocysts begin as unicellular bodies containing a single metrocyte and through asexual multiplication numerous metrocytes accumulate and the sarcocyst increases in size. As the sarcocyst matures, the small, rounded, noninfectious metrocytes give rise to crescent-shaped bodies called bradyzoites that are infections for the definitive host. Time required for maturation varies with the species and may take 2 months or more.
In species in which symptoms develop these typically occur 20–40 days after ingestion of sporocysts and during the subsequent migration of sporozoites through the body vessels. Acute lesions (oedema, hemorrhages and necrosis) develop in the affected tissues. The parasite has a predilection for skeletal muscle (myositis), cardiac muscle (petechial hemorrhages of cardiac muscle and serosae), and lymph nodes (oedema, necrosis and hemorrhage). These lesions are associated with maturation of second generation of meronts within the endothelial and subendothelials cells. Occasionally mononuclear infiltration or hyperemia has been observed in the lamina propria of the small intestine. After the acute phase cysts may be found in various muscular tissues, generally without pathology.
Once the intermediate host is eaten by the definitive host such as a dog or human, the parasite undergoes sexual reproduction within the gut to create macrogamonts and microgamonts. Most definitive hosts do not show any clinical sign or symptoms. Fusion of a macrogamont and a microgamont creates a zygote which develops into an oocyst. The oocyst is passed through the faeces completing the life cycle.
A second life cycle has more recently been described whereby carnivores and omnivores pass the infectious stages in their faeces. Ingestion of this material may lead to successful infection of the ingesting animal.
Birds
Although sarcocysts were first reported in the muscles of birds by Kuhn in 1865 not until 1977 was the first life cycle involving a bird (Gallus gallus) and a carnivore (Canis familiaris) described by Munday et al.[2] In 1986 the first life cycle involving birds as both the definitive (Northern goshawk - Accipiter gentilis) and intermediate (Atlantic canary - Serinus canaria) hosts was described by Cern and Kvasnovsk.[3]
Taxonomy
The taxonomy of this genus and its relationship to other protozoal genera is currently under investigation.
Related genera include: Besnoitia, Caryospora, Cystoisospora, Frenkelia, Isospora, Hammondia, Hyaloklossia, Lankesterella, Neospora and Toxoplasma.
Sarcocystis is the largest genus within the family Sarcocystidae and consists of species which infecting a range of animals including mammals, birds and reptiles. Frenkelia, another genus within this family, consists of parasites that use rodents as intermediate hosts and birds of prey as definitive hosts.
It appears that Besnoitia, Hammondia, Neospora and Toxoplasma form a single clade. Within this clade Toxoplasma and Neospora appear to be sister clades. Isospora also appear to belong to this clade and this clade is a sister to Sarcocystis. Frenkelia appears to be very closely related to Sarcocystis.
Several molecular studies have suggested that Frenkelia is actually a species of Sarcocystis. This genus was distinguished from Sarcocystis on the basis of its tendency to encyst within the brain rather than within muscle. This distinction may not be taxonomically valid.
Within the genus a number of clades have been identified. These include one that contains S. dispersa, S. lacertae, S. mucosa, S. muris, S. neurona and S. rodentifelis.[4] Frenkelia also groups with this clade.
Evolution
These protozoa are mostly found in mammals. They do not appear to infect mammals of the superorder Afrotheria and infect only two species of the Xenarthra. Because of this pattern the genus may have evolved in the Northern hemisphere from a preexisting protozoan species that infected mammals.
Alternatively because a number of Australian marsupials are also infected by this genus, marsupials may have been the original hosts of this genus and the parasites were spread to the Northern hemisphere by birds.
A third possibility is that the genus originally infected birds and was spread world wide by these hosts.
A final possibility because of the existence of life cycles where both the intermediate and final hosts are reptiles, the genus may have originated in reptiles and spread from there to other genera.
The resolution of this question awaits the outcome of further molecular studies.
Clinical: Human
Infection with this parasite is known as sarcosporidiosis. Because of initial confusion over the taxonomy of this parasite it was originally referred to as Isospora hominis. The older literature may refer to this organism.
Epidemiology
Human infection is considered rare with less than one hundred published cases of invasive disease (approximately 46 cases reported by 1990). These figures represent a gross underestimate of the human burden of disease.
The extremes of age reported to date are a 26-day-old infant and a 75-year-old man. Infectons have been reported from Africa, Europe (Germany, Spain and Poland), the United States (California), Central and South America, China, India, Tibet, Malaysia and Southeast Asia.
Stool examinations in Thai laborers showed that sarcocystis infection had a prevalence of ~23%. Virtually all cases appeared to be asymptomatic which probably explains the lack of recognition. A study of 100 human tongues obtained at post mortum in Malaya revealed an infection rate of 21%. There was no sex difference and the age range was 16 to 57 years (mean 37.7 years).[5]
History
The first report of human infection was by Lindemannl in 1868. Although several additional reports were subsequently published, these early descriptions were not considered definitive. The first generally agreed definitive description of this disease was published in 1894 by Baraban and Saint-Remy.[6] This species was named by Rivolta after Lindemannl in 1898.
The invasive forms were considered to belong to a single species - S. lindemanni - and the intestinal form due to S. hominis (from undercooked beef) or S. suihominis (from undercooked pork). The description of S. lindemanni has since been considered to be unsatisfactory and has been declared a nomem nudum (a name without a recognised species). Two species currently considered to be capable of causing human infection: S. bovihominis (S. hominis) and S. suihominis.
Route of infection
Infection occurs when undercooked meat is ingested. The incubation period is 9–39 days. Human outbreaks have occurred in Europe. Rats are a known carrier.
It has been suggested that contaminated water may be able to cause infection but this presently remains a theoretical possibility.
Pathology
The pathology is of two types: a rare invasive form with vasculitis and myositis and an intestinal form that presents with nausea, abdominal pain, and diarrhea. While normally mild and lasting under 48 hours, the intestinal form may occasionally be severe or even life-threatening. The invasive form may involve a wide variety of tissues including lymph nodes, muscles and the larynx.
Clinical features
In volunteer studies with infected beef symptoms appeared 3–6 hours after eating. These included anorexia, nausea, abdominal pain, distension, diarrhea, vomiting, dyspnoea and tachycardia. All symptoms were transient and lasted about 36 hours. In a second series symptoms - abdominal pain, distension, watery diarrhea and eosinophilia - appeared at 1 week and resolved after 3 weeks.
Clinical cases have been associated with acute fever, myalgias, bronchospasm, pruritic rashes, lymphadenopathy, subcutaneous nodules associated with eosinophilia, elevated erythrocyte sedimentation rate and elevated creatinine kinase levels. Symptoms may last as long as five years. Segmental necrotizing enteritis has been reported on one occasion.
Diagnosis
Definitive diagnosis by biopsy of an infected muscle. Sarcocysts are identifiable with hematoxylin and eosin. The PAS stain may be helpful but variable uptake of stain is common. Along with the sarcocysts inflammatory cells may be found. Other findings include myositis, myonecrosis, perivascular and interstitial inflammation, vasculitis and eosinophilic myositis.
Treatment
Because infection is rarely symptomatic, treatment is rarely required. There have been no published trials so treatment remains empirical. Agents that have been used include albendazole, metronidazole and cotrimoxazole for myositis. Corticosteroids have also been used for symptomatic relief.
Amprolium and salinomycin was effective in preventing severe illness and death in experimentally infected calves and lambs. These agents have not been tried in humans to date.
Prevention
Infection can be prevented by cooking the meat before eating. Alternatively freezing the meat at -5C for several days before ingestion will kill the sporocysts.
Clinical: Non human
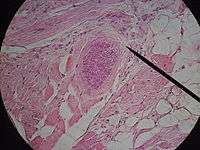
Four recognised species infect cattle: S. bovifelis, S. bovihominis (S. hominis) S. cruzi (S. bovicanis) and S. hirsuta. S. cruzi is the only species known to be pathogenic in cattle. A number of clinical syndromes have been reported in connection with this parasite: eosinophilic myositis; abortions, stillbirths and deaths in pregnant cows; two cases of necrotic encephalitis in heifers have also been reported. Typical clinical signs of acute bovine sarcocystosis are: anorexia, pyrexia (42C or more), anemia, cachexia, enlarged palpable lymph nodes, excessive salivation and loss of hair at the tip of the tail.
Sheep may be infected by four recognized species of Sarcocystis: S. arieticanis and S. tenella (S. ovicanis) are pathogenic; S. gigantea (S. ovifelis) and S. medusiformis are non-pathogenic. Infection with these parasites is common in the US with over 80% of sheep examined showing evidence of infection.[7] S. arieticanis and S. tenella both produce extra intestinal disease. Anemia, anorexia, ataxia, and abortions are the chief clinical signs. Myositis with flaccid paralysis has been reported as a consequence of infection. Ovine protozoan myeloencephalitis is a recognised syndrome that may occur in outbreaks. The usual pathological findings in such cases are multifocal spinal cord white matter oedema and necrosis, glial nodules and mild to moderate nonsuppurative encephalomyelitis. The diagnosis may be established finding protozoan bodies (12.7-23.0 micrometres) that stain immunocytochemically for Sarcocystis epitopes.
Four recognised species infect pigs: S. medusiformis, S. meischeriana (S. suicanis), S. porcifelis and S. suihominis. S. porcifelis is pathogenic for pigs causing diarrhea, myositis and lameness.
Five species infect horses: S. asinus, S. bertrami, S. equicanis, S. fayeri and S. neurona (S. falcatula). All utilize canids as definitive hosts: transplacental infection has also been reported. S. neurona causes equine protozoal myeloencephalitis. Exposure to this parasite appears to be common in the United States with serological surveys indicating that 50-60% percent of all horses in the Midwest United States have been exposed to it. Clinical signs include gait abnormalities including ataxia, knuckling and crossing over. Muscle atrophy, usually unilateral, may occur. The lesions are typically focal. Brain-stem involvement is common. Depression, weakness, head tilt and dysphagia also occur.
S. fayeri may cause myositis in horses.
Fatal infection of an alpaca (Lama pacos) with an unnamed species has been reported. Findings included disseminated eosinophilic myositis, abortion and haemoabdomen. The myositis was associated with hameorrhage, necrosis and degeration.[8] Infection by S. tilopodi of muscle tissue in the Guanaco has been reported.[9]
S. hemionilatrantis infects mule deer. Death from experimental inoculation have been reported.
These parasites can also infect birds producing three different clinical forms: an acute pulmonary disease, muscular disease and neurological disease. Symptoms include lethargy, shortness of breath, tail bobbing, yellow tinted droppings and sudden death.
The presence of the cysts in the muscle of wild birds is known as "rice breast".
Incidence in animals
Infection with Sarcocystis is common. Rates in pigs vary: 18% in Iowa,[10] 27% in the Philippines,[11] 43% in Spain,[12] 57% in Uruguay,[13] and 68% in India [14] The infection rate in sheep is commonly above 90%.[12][15][16][17] Camels have a similarly high incidence of infection.[15][17] Rates above 80% are known in cattle and goats.[15][16] The incidence in water buffaloes, yak and hainag exceeds 80%[15][17] while the incidence in horses, donkeys and chickens is lower.[16][17]
Diagnosis
There are several other genera of heteroxenous and cyst-forming coccidia[18] including Besnoitia, Cystoisospora, Frenkelia, Hammondia, Neospora and Toxoplasma.[19] Related but monoxenous spore forming genera include Isospora. Differentiating these genera from Sarcocystis in diagnostic material may be difficult without immunochemical stains.
The diagnosis is usually made post mortem by examination of the skeletal muscle. In some species the cysts may be visible to the naked eye (ducks, mice, rabbits and sheep) but in most microscopic examination is required. Ante mortem diagnosis may be made with the use of dermal sensitivity testing or complement fixation tests. Muscle biopsy is also diagnostic but this is much less commonly used.
Oocysts with two sporocysts or individual sporocysts in human feces are diagnostic of intestinal infection. These first appear 14 to 18 days after ingesting beef (S. hominis), and 11 to 13 days after ingesting pork (S. suihominis). Flotation based on high-density solutions incorporating sodium chloride, cesium chloride, zinc sulfate, sucrose, Percoll, Ficoll-Hypaque or other such density gradient media is preferred to formalin-ethyl acetate or other sedimentation methods. Sporocysts of S. hominis average 9.3 by 14.7 micrometres and those of S. suihominis average 10.5 by 13.5 micrometres. Because of the overlap in size, size alone is not reliable as a diagnostic criterion of the species. Confirmatory staining with the periodic acid-Schiff (PAS) can be performed as the walls stain positively. PCR amplification of the rRNA may also be used.
The walls of the sarcocyst may be helpful in species diagnosis with 24 wall types identified in 62 species. S. hominis and S. suihominis both have walls of type 10. The wall of S. hominis is up to 6 micrometres thick and appears radially striated from villar protrusions up to 7 micrometres long. Its bradyzoites are 7 to 9 micrometres long. The wall of S. suihominis is 4 to 9 micrometres thick, with villar protrusions up to 13 micrometres long. Its bradyzoites are 15 micrometres long.
Treatment
Current treatments are not entirely satisfactory. Amprolium (100 mg/kg, sid for 30 days), fed prophylactically, reduced illness in cattle inoculated with S. cruzi. Prophylactic administration of amprolium or salinomycin also protected experimentally infected sheep.
In horses treatment has been confined to dihydrofolate reductase inhibitors such as the sulfonamides and pyrimethamine. Sulfadiazine (20 mg/kg orally) once or twice a day is a commonly used. Infected horses should also be placed on pyrimethamine at the dose of 1.0 mg/kg given once a day orally for 120 days or longer. Diclazuril and Toltrazuril and other coccidiostats are being evaluated to treat EPM.
Vaccination
No vaccines are currently known. Experimentally inoculated pigs appear to develop a persistent immunity so a vaccine may be possible.
Host-parasite relations
The parasite's life cycle typically involves a predator and a prey animal. A single species may infect multiple prey or predator animals. In at least 56 species definitive and intermediate hosts are known. Many species are named after their recognised hosts.
A listing of the known host-parasite relations can be found on the page Sarcocystis: Host-parasite relations.
Genome
S. neurona is the subject of an ongoing (as of September 2011) genome project.
Notes
Hoareosporidium is now considered a synonym of Sarcocystis.
The original type species was Sarcocystis miescheriana. Its description has since been considered less than satisfactory and S. muris has been proposed as the type species.
S. turdi may not be a valid species.
Isospora bigemina has been reclassified as Sarcocystis bigemina.
Isospora hominis has been reclassified as Sarcocystis hominis.
S. bovihominis is a synonym of S. hominis.
S. cruzi is a synonym of S. bovicanis.
S. gigantea is a synonym of S. ovifelis.
S. hirsuta is a synonym of S. bovifelis.
S. idahoensis and S. roudabushi may be the same species.
S. miescheriana is a synonym of S. suicanis
S. neurona is a junior synonym of S. falcatula. S. neuroma is the more commonly used name for this species.
S. poephagi may be the same species as S. hirsuta.
S. poephagicanis may be the same species as S. cruzi.
S. tenella is a synonym of S. ovicanis.
See also
References
- ↑ Miescher F. (1843) Ueber eigenthiimliche Schlauche in den Muskeln einer Hausmaus. Ber. u.d. Verhandl. Naturf. Ges. Basel 5: 198-202
- ↑ Munday B.L, Humphrey J.D.,Ki1a V.(1977) Pathogy and produced by, preva1ence of, and probab1e life-cycle of a species of Sarcocystis in the domestic fowl. Avian Vis. 21: 697-703
- ↑ Cern Z & Kvasnovsk Z (1986) Life cycle involving bird-bird relation in sarcocystis coccidia with the description of Sarcocystis accipitris sp.n. Folia parasitologica 33, 305-309
- ↑ Elsheikha H.M., Lacher D.W., Mansfield L.S. (2005) Phylogenetic relationships of Sarcocystis neurona of horses and opossums to other cyst-forming coccidia deduced from SSU rRNA gene sequences. Parasitol. Res. 97(5):345-357
- ↑ Wong K.T., Pathmanathan R. (1992) High prevalence of human skeletal muscle sarcocystosis in south-east Asia. Trans. R. Soc. Trop. Med. Hyg. 86(6):631-632.
- ↑ Baraban M. Le and Saint-Remy M. G.(1894) Sur un cas de tubes psorospermiques observes chez 1' homme. Compt. Rend. Soc. de Biol. 46: 231-203
- ↑ Dubey J.P., Lindsay D.S., Speer C.A., Fayer R., Livingston C.W. Jr. (1988) Sarcocystis arieticanis and other Sarcocystis species in sheep in the United States. J. Parasitol. 74(6):1033-1038
- ↑ La Perle K.M., Silveria F., Anderson D.E., Blomme E.A. (1999) Dalmeny disease in an alpaca (Lama pacos): sarcocystosis, eosinophilic myositis and abortion. J. Comp. Pathol. 121(3):287-293
- ↑ C. Michael Hogan. 2008. Guanaco: Lama guanicoe, GlobalTwitcher.com, ed. N. Strömberg
- ↑ Dubey J.P., Powell E.C. (1994) Prevalence of Sarcocystis in sows from Iowa. Vet. Parasitol. 52(1-2):151-155
- ↑ Claveria F.G., De La Peña C., Cruz-Flores M.J., Saleque A., Bhatia B.B. Freyre A., Chifflet L., Mendez J. (2001) Sarcocystis miescheriana infection in domestic pigs (Sus scrofa) in the Philippines. J. Parasitol. 87(4):938-939.
- 1 2 Pereira A., Bermejo M. (1988) Prevalence of Sarcocystis cysts in pigs and sheep in Spain. Vet. Parasitol. 27(3-4):353-355
- ↑ Freyre A, Chifflet L., Mendez J. Sarcosporidian infection in pigs in Uruguay. Vet. Parasitol. 41(1-2):167-171
- ↑ Saleque A., Bhatia B.B. (1991)Prevalence of Sarcocystis in domestic pigs in India. Vet. Parasitol. 40(1-2):151-153
- 1 2 3 4 Latif B.M., Al-Delemi J.K., Mohammed B.S., Al-Bayati S.M., Al-Amiry A.M. (1999)Prevalence of Sarcocystis spp. in meat-producing animals in Iraq. Vet. Parasitol. 84(1-2):85-90
- 1 2 3 Woldemeskel M., Gebreab F. (1996) Prevalence of sarcocysts in livestock of northwest Ethiopia. Zentralbl Veterinarmed B. 43(1):55-58.
- 1 2 3 4 Fukuyo M., Battsetseg G., Byambaa B. (2002) Prevalence of Sarcocystis infection in meat-producing animals in Mongolia. Southeast Asian J. Trop. Med. Public Health. 33(3):490-495.
- ↑ "Coccidia Parasite Life Cycle". Retrieved 12 May 2014.
- ↑ Gregarines, Haemogregarines, Coccidia, Plasmodia, and Haemoproteids. 216: Elsevier. 1977. p. 580. ISBN 0323163254.
External links
- Sarcocystis at the US National Library of Medicine Medical Subject Headings (MeSH)
- Sarcocystis genome project
- Review: http://cmr.asm.org/cgi/content/full/17/4/894
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||