Rho family of GTPases
The Rho family of GTPases is a family of small (~21 kDa) signaling G proteins, and is a subfamily of the Ras superfamily. The members of the Rho GTPase family have been shown to regulate many aspects of intracellular actin dynamics, and are found in all eukaryotic kingdoms, including yeasts and some plants. Three members of the family have been studied in detail: Cdc42, Rac1, and RhoA. All G proteins are "molecular switches", and Rho proteins play a role in organelle development, cytoskeletal dynamics, cell movement, and other common cellular functions.[1][2]
History
Identification of the Rho family of GTPases began in the mid-1980s. The first identified Rho member was RhoA, isolated serendipitously in 1985 from a low stringency cDNA screening.[3] Rac1 and Rac2 were identified next, in 1989[4] followed by Cdc42 in 1990.[5] Eight additional mammalian Rho members were identified from biological screenings until the late 1990s, a turning point in biology where availability of complete genome sequences allowed full identification of gene families. All eukaryote cells contain Rho GTPase (ranging from 6 in yeast to 20 in mammals). In mammals, the Rho family is thus made of 20 members distributed in 8 subfamilies: Rho, Rnd, RhoD/F, RhoH, Rac, Cdc42, RhoU/V and RhoBTB.[1]
As early as 1990, Paterson et al. began expressing activated Rho protein in Swiss 3T3 fibroblasts.[6]
By the mid-1990s, Rho proteins had been observed to affect the formation of cellular projections ("processes") in fibroblasts. In a 1998 review article, Alan Hall compiled evidence showing that not only do fibroblasts form processes upon Rho activation, but so do virtually all eukaryotic cells.[7]
A 2006 review article by Bement et al. explored the significance of spatial zones of Rho activation.[8]
Categorization
The Rho family of GTPases belong to the Ras superfamily of proteins, which consists of over 150 varieties in mammals. Rho proteins sometimes denote some members of the Rho family (RhoA, RhoB, and RhoC), and sometimes refers to all members of the family. This article is about the family as a whole.
In mammals, the Rho family contains 22 members.<ref name="Ridley[9] Almost all research involves the three most common members of the Rho family: Cdc42, Rac1 and RhoA.
| Rho family member | Action on actin filaments |
|---|---|
| Cdc42 | affects filopodia |
| Rac1 | affects lamellipodia |
| RhoA | affects stress fibres |
These 22 mammalian members are subdivided in the Rac subfamily (Rac1, Rac2, Rac3, and RhoG), Cdc42 subfamily (Cdc42, TC10, TCL, Chip, and Wrch-1), RhoA subfamlly (RhoA, RhoB, and RhoC) and other Rho GTPases (RhoE/Rnd3, RhoH/TTF, Rif, RhoBTB1, RhoBTB2, Miro-1, Miro-2, RhoD, Rnd1, and Rnd2).[9]
| Subclass | Cytoskeletal effect | Rho family members |
|---|---|---|
| Cdc42 subclass | filopodia | Cdc42 |
| RhoQ (TC10) | ||
| RhoJ (TCL) | ||
| RhoUV subclass | filopodia and lamellipodia | RhoU (Wrch) |
| RhoV (Chp) | ||
| Rac | lamellipodia | Rac1 |
| Rac2 | ||
| Rac3 | ||
| RhoG | ||
| RhoBTB | protein stability | RhoBTB1 |
| RhoBTB2 | ||
| RhoBTB3 | ||
| RhoH | Rac agonist? | RhoH |
| Rho (subclass) | ↑stress fibres and ↑focal adhesions | RhoA |
| RhoB | ||
| RhoC | ||
| Rnd | ↓stress fibres and ↓focal adhesions | Rnd1 |
| Rnd2 | ||
| Rnd3 (RhoE) | ||
| RhoF | Vesicle transport, filopodia | RhoD |
| RhoF (Rif) |
Regulators
Three general classes of regulators of Rho protein signaling have been identified: guanine nucleotide exchange factor (GEFs), GTPase-activating proteins (GAPs) and guanine nucleotide dissociation inhibitors (GDIs).[10] GEFs activate Rho proteins by catalyzing the exchange of GDP for GTP. GAPs control the ability of the GTPase to hydrolyze GTP to GDP, controlling the natural rate of movement from the active conformation to the inactive conformation. GDI proteins form a large complex with the Rho protein, helping to prevent diffusion within the membrane and into the cytosol and thus acting as an anchor and allowing tight spatial control of Rho activation.[10]
Recent work has unveiled important additional regulatory mechanisms: microRNAs regulate post-transcriptional processing of Rho GTPase-encoding mRNAs; palmitoylation and nuclear targeting affect intracellular distribution; post-translational phosphorylation, transglutamination and AMPylation modulate Rho GTPase signaling; and ubiquitination controls Rho GTPase protein stability and turnover. These modes of regulation add to the complexity of the Rho GTPase signaling network and allow precise spatiotemporal control of individual Rho GTPases.[11]
Effectors
Each Rho protein affects numerous proteins downstream, all of which having roles in various cell processes. Over 60 targets of the three common Rho GTPases have been found.[12] Two molecules that directly stimulate actin polymerization are the Arp2/3 proteins and the Diaphanous-related formins.[13]
| GTPase | Effector[2][13] |
|---|---|
| RhoA | Cit, Cnksr1, Diaph1, Diaph2, DgkQ, FlnA, KcnA2, Ktn1, Rtkn1, Rtkn2, Rhpn1, Rhpn2, Itpr1, PlcG1, PI-5-p5K, Pld1, Pkn1, Pkn2, Rock1, Rock2, PrkcA, Ppp1r12A |
| Rac1 | Sra1, IRSp53, PAK1, PAK2, PAK3 |
| Cdc42 | Wiskott-Aldrich syndrome protein, N-WASP, IRSp53, Dia2, Dia3, ROCK1, ROCK2 |
Functions
Rho/Rac proteins are involved in a wide variety of cellular functions such as cell polarity, vesicular trafficking, the cell cycle and transcriptomal dynamics.[2]
Morphology
Animal cells form many different shapes based on their function and location in the body. Rho proteins help cells regulate changes in shape throughout their life-cycle. Before cells can undergo key processes such as budding, mitosis, or locomotion, it must have some manner of cell polarity.
One example of Rho GTPases' role in cell polarity is seen in the much-studied yeast cell. Before the cell can bud, Cdc42 is used to locate the region of the cell's membrane that will begin to bulge into the new cell. When Cdc42 is removed from the cell, the outgrowths still form, but do so in an unorganized manner.[12]
One of the most obvious changes to cell morphology controlled by Rho proteins is the formation of lamellipodia and filopodia, projecting processes that look like "fingers" or "feet" and often propel cells or growth cones across surfaces. Virtually all eukaryotic cells form such processes upon Rho activation.[7] Fibroblasts such as Swiss 3T3 cells are often used to study these phenomena.
Study techniques
Much of what is known about cellular morphology changes and the effects of Rho proteins comes from the creation of a constitutively active mutated form of the protein. Mutation of a key amino acid can alter the conformation of the entire protein, causing it to permanently adopt a conformation that resembles the GTP-bound state.[6] This protein cannot be inactivated normally, through GTP hydrolysis, and is thus "stuck on". When a Rho protein activated in this manner is expressed in 3T3 cells, morphological changes such as contractions and filopodia formation ensue.[6]
Because Rho proteins are G-proteins and plasma membrane bound, their location can be easily controlled. In each situation, whether it be wound healing, cytokinesis, or budding, the location of the Rho activation can be imaged and identified. For example, if a circular hole is inflicted in a spherical cell, Cdc42 and other active Rhos are seen in highest concentration around the circumference of the circular injury.[8] One method of maintaining the spatial zones of activation is through anchoring to the actin cytoskeleton, keeping the membrane-bound protein from diffusing away from the region where it is most needed.[8] Another method of maintenance is through the formation of a large complex that is resistant to diffusion and more rigidly bound to the membrane than the Rho itself.[8]
Morphogenesis of dendritic spines
The morphogenesis of dendritic spines is critical to the induction of long-term potentiation (LTP).[14][15] The morphology of the spine depends on the states of actin, either in globular (G-actin) or filamentous (F-actin) forms. The role of Rho family of GTPases and its effects in the stability of actin and spine motility[16] has important implications for memory. If the dendritic spine is the basic unit of information storage, then the spine's ability to extend and retract spontaneously must be constrained. If not, information may be lost. Rho family of GTPases makes significant contributions to the process that stimulates actin polymerization, which in turn increases the size and shape of the spine.[17] Large spines are more stable than smaller ones and may be resistant to modification by additional synaptic activity.[18] Because changes in the shape and size of dendritic spines are correlated with the strength of excitatory synaptic connections and heavily depend on remodeling of its underlying actin cytoskeleton,[19] the specific mechanisms of actin regulation, and therefore the Rho family of GTPases, are integral to the formation, maturation, and plasticity of dendritic spines and to learning and memory.
RhoA pathway
One of the major Rho GTPases involved in spine morphogenesis is RhoA, a protein that also modulates the regulation and timing of cell division. In the context of activity in neurons, RhoA is activated in the following manner: once calcium has entered a cell through NMDA receptors, it binds to calmodulin and activates CaMKII, which leads to the activation of RhoA.[17] The activation of the RhoA protein will activate ROCK, a RhoA kinase, which leads to the stimulation of LIM kinase, which in turn inhibits the protein cofilin. Cofilin's function is to reorganize the actin cytoskeleton of a cell; namely, it depolymerizes actin segments and thus inhibits the growth of growth cones and the repair of axons.[20]
A study conducted by Murakoshi et al. in 2011 implicated the Rho GTPases RhoA and Cdc42 in dendritic spine morphogenesis. Both GTPases were quickly activated in single dendritic spines of pyramidal neurons in the CA1 region of the rat hippocampus during structural plasticity brought on by long-term potentiation stimuli. Concurrent RhoA and Cdc42 activation led to a transient increase in spine growth of up to 300% for five minutes, which decayed into a smaller but sustained growth for thirty minutes.[17] The activation of RhoA diffused around the vicinity of the spine undergoing stimulation, and it was determined that RhoA is necessary for the transient phase and most likely the sustained phase as well of spine growth.
Cdc42 pathway
Cdc42 has been implicated in many different functions including dendritic growth, branching, and branch stability.[21] Calcium influx into the cell through NMDA receptors binds to calmodulin and activates the Ca2+/calmodulin-dependent protein kinases II (CaMKII). In turn, CaMKII is activated and this activates Cdc42, after which no feedback signaling occurs upstream to calcium and CaMKII. If tagged with monomeric-enhanced green fluorescent protein, one can see that the activation of Cdc42 is limited to just the stimulated spine of a dendrite. This is because the molecule is continuously activated during plasticity and immediately inactivates after diffusing out of the spine. Despite its compartmentalized activity, Cdc42 is still mobile out of the stimulated spine, just like RhoA. Cdc42 activates PAK, which is a protein kinase that specifically phosphorylates and, therefore, inactivates ADF/cofilin.[22] Inactivation of cofilin leads to increased actin polymerization and expansion of the spine's volume. Activation of Cdc42 is required for this increase in spinal volume to be sustained.
Observed changes in structural plasticity
Murakoshi, Wang, and Yasuda (2011) examined the effects of Rho GTPase activation on the structural plasticity of single dendritic spines elucidating differences between the transient and sustained phases.[17]
Transient changes in structural plasticity
Applying a low-frequency train of two-photon glutamate uncaging in a single dendritic spine can elicit rapid activation of both RhoA and Cdc42. During the next two minutes, the volume of the stimulated spine can expand to 300 percent of its original size. However, this change in spine morphology is only temporary; the volume of the spine decreases after five minutes. Administration of C3 transferase, a Rho inhibitor, or glycyl-H1152, a Rock inhibitor, inhibits the transient expansion of the spine, indicating that activation of the Rho-Rock pathway is required in some way for this process.[17]
Sustained changes in structural plasticity
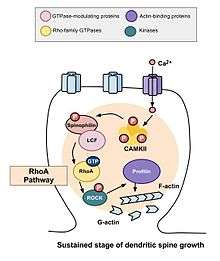
After the transient changes described above take place, the spine's volume decreases until it is elevated by 70 to 80 percent of the original volume. This sustained change in structural plasticity will last about thirty minutes. Once again, administration of C3 transferase and Glycyl-H1152 suppressed this growth, suggesting that the Rho-Rock pathway is necessary for more persistent increases in spinal volume. In addition, administration of the Cdc42 binding domain of Wasp or inhibitor targeting Pak1 activation-3 (IPA3) decreases this sustained growth in volume, demonstrating that the Cdc42-Pak pathway is needed for this growth in spinal volume as well. This is important because sustained changes in structural plasticity may provide a mechanism for the encoding, maintenance, and retrieval of memories. The observations made may suggest that Rho GTPases are necessary for these processes.[17]
Movement
In addition to the formation of lamellipodia and filopodia, intracellular concentration and cross-talk between different Rho proteins drives the extensions and contractions that cause cellular locomotion. Sakumura et al. proposed a model based on differential equations that helps explain the activity of Rho proteins and their relationship to motion. This model encompassed the three proteins Cdc42, RhoA, and Rac. Cdc42 was assumed to encourage filopodia elongation and block actin depolymerization. RhoA was considered to encourage actin retraction. Rac was treated to encourage lamellipodia extension but block actin depolymerization. These three proteins, although significantly simplified, covered the key steps in cellular locomotion. Through various mathematical techniques, solutions to the differential equations that described various regions of activity based on intracellular activity were found. The paper concludes by showing that the model predicts that there are a few threshold concentrations that cause interesting effects on the activity of the cell. Below a certain concentration, there is very little activity, causing no extension of the arms and feet of the cell. Above a certain concentration, the Rho protein causes a sinusoidal oscillation much like the extensions and contractions of the lamellipodia and filopodia. In essence, this model predicts that increasing the intracellular concentration of these three key active Rho proteins causes an out-of-phase activity of the cell, resulting in extensions and contractions that are also out of phase.[23]
Wound healing
One example of behavior that is modulated by Rho GTPase proteins is in the healing of wounds. Wounds heal differently between young chicks and adult chickens. In young chicks, wounds heal by contraction, much like a draw-string being pulled to close a bag. In older chickens, cells crawl across the wound through locomotion. The actin formation required to close the wounds in young chicks is controlled by Rho GTPase proteins, since, after injection of a bacterial exoenzyme used to block rho and rac activity, the actin polymers do not form, and thus the healing completely fails.[24]
Phagocytosis
Another cellular behavior that is affected by rho proteins is phagocytosis. As with most other types of cell membrane modulation, phagocytosis requires the actin cytoskeleton in order to engulf other items. The actin filaments control the formation of the phagocytic cup, and active Rac1 and Cdc42 have been implicated in this signaling cascade.[25]
Mitosis
Yet another major aspect of cellular behavior that is thought to include rho protein signaling is mitosis. While rho GTPase activity was thought for years to be restricted to actin polymerization and therefore to cytokinesis, which occurs after mitosis, new evidence has arisen that shows some activity in microtubule formation and the process of mitosis itself. This topic is still debated, and there is evidence both for and against for the importance of rho in mitosis.[26]
Applications
Nervous system regeneration
Because of their implications in cellular motility and shape, Rho proteins became a clear target in the study of the growth cones that form during axonal generation and regeneration in the nervous system. Rho proteins may be a potential target for delivery into spinal cord lesions after traumatic injury. Following injury to the spinal cord, the extracellular space becomes inhibitory to the natural efforts neurons undergo to regenerate.
These natural efforts include the formation of a growth cone at the proximal end of an injured axon. Newly formed growth cones subsequently attempt to "crawl" across the lesion. These are sensitive to chemical cues in the extracellular environment. One of the many inhibitory cues includes chondroitin sulfate proteoglycans (CSPGs). Neurons growing in culture become more able to cross regions of substrate coated with CSPG after expression of constitutively active Cdc42 or Rac1[27] or expression of a dominant negative form (inhibition) of RhoA. This is partly due to the exogenous Rho proteins driving cellular locomotion despite the extracellular cues promoting apoptosis and growth cone collapse. Intracellular modulation of Rho proteins has thus become of interest in research aimed at spinal cord regeneration.
Mental retardation
Dysfunction of Rho proteins has also been implicated in mental retardation. Mental retardation in some cases involves malformation of the dendritic spines, which form the post-synaptic connections between neurons. The misshapen dendritic spines can result from modulation of rho protein signaling. After the cloning of various genes implicated in X-linked mental retardation, three genes that have effects on Rho signaling were identified, including oligophrenin-1 (a GAP protein that stimulates GTPase activity of Rac1, Cdc42, and RhoA), PAK3 (involved with the effects of Rac and Cdc42 on the actin cytoskeleton) and αPIX (a GEF that helps activate Rac1 and Cdc42).[28] Because of the effect of Rho signaling on the actin cytoskeleton, genetic malfunctions of a rho protein could explain the irregular morphology of neuronal dendrites seen in some cases of mental retardation.
Cancer
After finding that Ras proteins are mutated in 30% of human cancers, it was suspected that mutated Rho proteins might also be involved in cancer reproduction.[10] However, as of August 2007, no oncogenic mutations have been found in Rho proteins, and only one has been found to be genetically altered.[10] To explain the role of Rho pathways without mutation, researchers have now turned to the regulators of rho activity and the levels of expression of the Rho proteins for answers.
One way to explain altered signaling in the absence of mutation is through increased expression. Overexpression of RhoA, RhoB, RhoC, Rac1, Rac2, Rac3, RhoE, RhoG, RhoH, and Cdc42 has been shown in multiple types of cancer.[10] This increased presence of so many signaling molecules implies that these proteins promote the cellular functions that become overly active in cancerous cells.
A second target to explain the role of the Rho proteins in cancer is their regulatory proteins. Rho proteins are very tightly controlled by a wide variety of sources, and over 60 activators and 70 inactivators have been identified.[12] Multiple GAPs, GDIs, and GEFs have been shown to undergo overexpression, downregulation, or mutation in different types of cancer.[10] Once an upstream signal is changed, the activity of its targets downstream—i.e., the Rho proteins—will change in activity.
Ellenbroek et al. outlined a number of different effects of Rho activation in cancerous cells. First, in the initiation of the tumor modification of Rho activity can suppress apoptosis and therefore contribute to artificial cell longevity. After natural apoptosis is suppressed, abnormal tumor growth can be observed through the loss of polarity in which Rho proteins play an integral role. Next, the growing mass can invade across its normal boundaries through the alteration of adhesion proteins potentially caused by Rho proteins.[10] Finally, after inhibition of apoptosis, cell polarity and adhesion molecules, the cancerous mass is free to metastasize and spread to other regions of the body.
References
- 1 2 Boureux A, Vignal E, Faure S, Fort P. (2007). "Evolution of the Rho family of ras-like GTPases in eukaryotes". Mol Biol Evol 24 (1): 203–16. doi:10.1128/JB.00142-07. ISSN 0021-9193. PMC 1913362. PMID 17449622.
- 1 2 3 Bustelo XR, Sauzeau V, Berenjeno IM. (2007). "GTP-binding proteins of the Rho/Rac family: regulation, effectors and functions in vivo". BioEssays 29 (4): 356–370. doi:10.1002/bies.20558. PMC 1971132. PMID 17373658.
- ↑ Madaule P., Axel R. (1985). "A novel ras-related gene family". Cell 41 (1): 31–40. doi:10.1016/0092-8674(85)90058-3. PMID 3888408.
- ↑ Didsbury J, Weber RF, Bokoch GM, Evans T, Snyderman R. (1989). "Rac, a novel ras-related family of proteins that are botulinum toxin substrates". J Biol Chem 264 (28): 16378–82. ISSN 0021-9258. PMID 2674130.
- ↑ Munemitsu S, Innis M, Clark R, McCormick F, Ullrich A, Polakis P. (1990). "Molecular cloning and expression of a G25K cDNA, the human homolog of the yeast cell cycle gene CDC42". Mol Cell Biol 10 (11): 5977–82. ISSN 0270-7306. PMC 361395. PMID 2122236.
- 1 2 3 Paterson HF, Self AJ, Garrett MD, Just I, Aktories K, Hall A. (1990). "Microinjection of recombinant p21 rho induces rapid changes in cell morphology". J Cell Biol 111 (3): 1001–7. doi:10.1083/jcb.111.3.1001. PMC 2116288. PMID 2118140.
- 1 2 Hall A. (1998). "Rho GTPases and the actin cytoskeleton". Science 279 (5350): 509–14. doi:10.1126/science.279.5350.509. PMID 9438836.
- 1 2 3 4 Bement WM, Miller AL, von Dassow G. (2006). "Rho GTPase activity zones and transient contractile arrays". BioEssays 28 (10): 983–93. doi:10.1002/bies.20477. PMID 16998826.
- 1 2 Ridley, AJ; et al. (Oct 2006). "Rho GTPases and actin dynamics in membrane protrusions and vesicle trafficking". Trends Cell Biol 16 (10): 522–9. doi:10.1016/j.tcb.2006.08.006. PMID 16949823.
- 1 2 3 4 5 6 7 Ellenbroek S, Collard J. (2007). "RhoGTPases: functions and association with cancer". Clin Exp Metastasis 24 (8): 657–72. doi:10.1007/s10585-007-9119-1. PMID 18000759.
- ↑ Meng Liu, Yi Zheng (2012). "Rho GTPase regulation by miRNAs and covalent modifications". Trends in Cell Biology 22 (7): 367–373. doi:10.1016/j.tcb.2012.04.004. PMID 22572609.
- 1 2 3 Etienne-Manneville S, Hall A. (2002). "Rho GTPases in cell biology". Nature 420 (6916): 629–35. doi:10.1038/nature01148. PMID 12478284.
- 1 2 Ridley A. (2006). "Rho GTPases and actin dynamics in membrane protrusions and vesicle trafficking". Trends Cell Biol 16 (10): 522–9. doi:10.1016/j.tcb.2006.08.006. PMID 16949823.
- ↑ Kim, Chong-Hyun; Lisman (1999). "A role of actin filament in synaptic transmission and long-term potentiation". Journal of Neuroscience 19 (11): 4314–4324. PMID 10341235.
- ↑ Krucker, T.; Siggins, G. R.; Halpain, S. (23 May 2000). "Dynamic actin filaments are required for stable long-term potentiation (LTP) in area CA1 of the hippocampus". Proceedings of the National Academy of Sciences 97 (12): 6856–6861. doi:10.1073/pnas.100139797.
- ↑ Tashiro, Ayumu; Yuste, Rafael (1 July 2004). "Regulation of dendritic spine motility and stability by Rac1 and Rho kinase: evidence for two forms of spine motility". Molecular and Cellular Neuroscience 26 (3): 429–440. doi:10.1016/j.mcn.2004.04.001. PMID 15234347.
- 1 2 3 4 5 6 Murakoshi, Hideji; Wang, Hong; Yasuda, Ryohei (20 March 2011). "Local, persistent activation of Rho GTPases during plasticity of single dendritic spines". Nature 472 (7341): 100–104. doi:10.1038/nature09823. PMC 3105377. PMID 21423166.
- ↑ Kasai, Haruo; Matsuzaki, Masanori; Noguchi, Jun; Yasumatsu, Nobuaki; Nakahara, Hiroyuki (1 July 2003). "Structure–stability–function relationships of dendritic spines". Trends in Neurosciences 26 (7): 360–368. doi:10.1016/S0166-2236(03)00162-0. PMID 12850432.
- ↑ Hotulainen, P.; Hoogenraad, C. C. (10 May 2010). "Actin in dendritic spines: connecting dynamics to function". The Journal of Cell Biology 189 (4): 619–629. doi:10.1083/jcb.201003008. PMC 2872912. PMID 20457765.
- ↑ Kiss, C.; Li, J.; Szeles, A.; Gizatullin, R.Z.; Kashuba, V.I..; Lushnikova, T.L.; Protopopov, A.I.; Kelve, M.; Kiss, H.; Kholodnyuk, I.D.; Irnreh, S.; Klein, G.; Zabarovsky, E.R. (1 January 1997). "Assignment of the ARHA and GPX1 genes to human chromosome bands 3p21.3 by in situ hybridization and with somatic cell hybrids". Cytogenetic and Genome Research 79 (3–4): 228–230. doi:10.1159/000134729. PMID 9605859.
- ↑ Scott, Ethan K.; Reuter, J.; Luo, L. (11 April 2003). "Small GTPase Cdc42 is required for multiple aspects of dendritic morphogenesis". Journal of Neuroscience 28 (8): 3118–3123.
- ↑ Calabrese, Barbara; Wilson, M.S.; Halpain, S. (1 February 2006). "Development and Regulation of Dendritic Spine Synapses". Physiology 21 (1): 38–47. doi:10.1152/physiol.00042.2005.
- ↑ Sakumura Y, Tsukada Y, Yamamoto N, Ishii S. (2005). "A molecular model for axon guidance based on cross talk between rho GTPases". Biophys J 89 (2): 812–22. doi:10.1529/biophysj.104.055624. PMC 1366631. PMID 15923236.
- ↑ Brock J, Midwinter K, Lewis J, Martin P. (1996). "Healing of incisional wound in the embryonic chick wing bud: characterization of the actin purse-string and demonstration of a requirement for Rho activation". J Cell Biol 135 (4): 1097–107. doi:10.1083/jcb.135.4.1097. PMC 2133375. PMID 8922389.
- ↑ Niedergang F, Chavrier P. (2005). "Regulation of phagocytosis by Rho GTPases". Curr Top Microbiol Immunol. Current Topics in Microbiology and Immunology 291: 43–60. doi:10.1007/3-540-27511-8_4. ISBN 3-540-23865-4. PMID 15981459.
- ↑ Narumiya S, Yasuda S. (2006). "Rho GTPases in animal cell mitosis". Curr Opin Cell Biol 18 (2): 199–205. doi:10.1016/j.ceb.2006.02.002. PMID 16487696.
- ↑ Jain A, Brady-Kalnay SM, Bellamkonda RV. (2004). "Modulation of Rho GTPase activity alleviates chondroitin sulfate proteoglycan-dependent inhibition of neurite extension". J Neurosci Res 77 (2): 299–307. doi:10.1002/jnr.20161. PMID 15211597.
- ↑ Ramakers GJ. (2002). "Rho proteins, mental retardation and the cellular basis of cognition". Trends Neurosci 25 (4): 191–9. doi:10.1016/S0166-2236(00)02118-4. PMID 11998687.
Several mutations in Rho proteins have been identified in large scale sequencing of cancers. These mutations are listed in the Catalogue of Somatic Mutations database (http://www.sanger.ac.uk/genetics/CGP/cosmic/). The functional consequences of these mutations are unknown.
See also
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||||||