Prodeinotherium
| Prodeinotherium Temporal range: 20.5–15.5 Ma | |
|---|---|
 | |
| Prodeinotherium bavaricum | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Proboscidea |
| Family: | †Deinotheriidae |
| Subfamily: | †Deinotheriinae |
| Genus: | †Prodeinotherium Ehik, 1930 |
| Species[1] | |
| Synonyms[1][3] | |
|
Prodinotherium Ehik, 1930 P. bavaricum
P. pentapotamiae
P. hobleyi
| |
Prodeinotherium is an extinct representative of the family Deinotheriidae that lived in Africa, Europe, and Asia in the early and middle Miocene. Prodeinotherium was first named in 1930, meaning "before terrible beast", but soon after the only species in it, P. hungaricum, was reassigned to Deinotherium. During the 1970s however, the two genera were once again separated, with Prodeinotherium diagnosed to include Deinotherium bavaricum (=P. hungaricum), Deinotherium hobleyi, and Deinotherium pentapotamiae, which were separated based on geographic location. The three species are from Europe, Africa, and Asia respectively. However, because of usage of few characters to separate them, there may have been only one species, P. bavaricum, or many more species, including P. cuvieri, P. orlovii, and P. sinense.
Prodeinotherium is one of three genera of Deinotheriidae, the others being Chilgatherium from Africa, and Deinotherium from Europe, Africa and Asia. Chilgatherium preceded Prodeinotherium, while Deinotherium succeeded it. P. hobleyi was the first species of Prodeinotherium, and it migrated into Asia and Europe before evolving into P. pentapotamiae and then P. bavaricum. Prodeinotherium lived for the Early Miocene and Middle Miocene before being replaced by Deinotherium. The deinotheriids are an early branch of proboscideans, although more derived than Barytherium and Moeritherium.
All deinotheres were large animals that evolved to be even larger, and many features are shared throughout the group. Prodeinotherium and Deinotherium both had large downcurved tusks on the lower jaw, but none on the upper jaw. They may have had a tapir-like snout, muscular but not as long as an elephant's trunk. This could have been used to grasp food while the tusks moved branches out of the way. Prodeinotherium was slightly smaller than Deinotherium, yet much larger than more primitive proboscideans. All Prodeinotherium species were similar in size, ranging from 2.5 to 2.8 m (8.2 to 9.2 ft) tall and weighing about 3.1 to 4.3 t (490 to 680 st).
Description

Prodeinotherium was the size of the present Asian elephant, about 3–4 metres (9.8–13.1 ft) at the shoulders, but differing from elephants by lacking upper tusks and instead possessing downward-facing lower tusks.[4][5] In appearance and many characters it was like Deinotherium, but differed in being of smaller size, having shorter forelimbs, and also in various details in the shape and form of the teeth.[2] An potentially adult female specimen of P. bavaricum is estimated to be 2.47 metres (8.1 ft) tall and weight 3.1 tonnes (3.1 long tons; 3.4 short tons), while an adult male measured 2.78 metres (9.1 ft) tall and was about 4.3 tonnes (4.2 long tons; 4.7 short tons). The earliest species P. hobleyi was estimated at similar 2.7 metres (8.9 ft) tall and 4.0 tonnes (3.9 long tons; 4.4 short tons).[4] Prodeinotherium hobleyi was larger and more specialised than its Oligocene predecessor Chilgatherium. It flourished for several millions of years, before being replaced in the middle Miocene by the much larger Deinotherium.[2]
Prodeinotherium is distinguished from Deinotherium from multiple features, including: possessing a different dental formula of 003/103 and 0023/1023; M2-3 with an ornamentation; the rostrum turns down parallel to the mandibular symphysis; the rostrum and external nares narrow; the swelling of the preorbital is close to the orbit; the roof of the skull is longer and wider than in Deinotherium; the articulation between the neck vertebrae and skull is more upturned; the skeleton is graviportally adapted; the scapula has a prominent spine and a stout acromion and metacromion; and the carpal bones and tarsal bones are narrow, but not dolichopodous.[1]
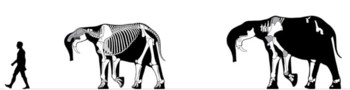
Deinotheres like Prodeinotherium have a muscle attachment for a trunk-like structure. However, a study in 2001 found that instead of an elephant-like trunk the appendage was more muscular and similar to a tapirs snout. They also found that within the evolution of Prodeinotherium, the pair "tongs" at the end of the trunk were lost.[5]
P. bavaricum is the most well-studied species of Prodeinotherium, with multiple features shared among all specimens, not necessarily to the exclusion of other species. Some of these features include "small size, generally simple dental structure, less enamel plication and crenulation and thus the valleys of the premolars are well separated, slender teeth, bicuspid mesial lophid in P3 (the cuspids are distinct but more compressed against each other than in P. hobleyi), and clear mesial projection (“preprotolophide”) in P3; sometimes is bicuspid." Other features noted earlier in 1957 include "the mesial lophid of P3 is well separated into two cuspids, the mesial projection of P3 is well developed and often bicuspid, and the base of the protoconid in P3 is longer than that of the metaconid." P. hobleyi differs in morphology from P. bavaricum mostly in these P3 characteristics.[6]
All deinothere mandibles have the same basic anatomy, with a downturned symphysis, and lower incisors. Most differences of deinothere genera are in the P3 tooth morphology and dimensions of the mandible and teeth. Measurements of the mandible have shown that the curve of the jaw is relative to the length of the jaw, a longer jaw and a stronger curve. A distinguishing feature of Prodeinotherium is that the area at the base of the curve in the jaw is flat, while there is a depression in all specimens of Deinotherium.[7]
Taxonomy and evolution

Prodeinotherium lived during the Early Miocene and Middle Miocene, about 19.0 to 18.0 million years ago.[6] Prodeinotherium likely evolved from Chilgatherium, or the common ancestor of the two genera.[2] The earliest remains of Prodeinotherium come from Kenya, where two deposits preserving the genus date to 22.5 and 19.5 million years ago according to one 1978 study. The same study found that fossils from Uganda date to 20.0 may. However, more recent studies (from 1988, 1991 and 2002) find that the deposits date to >17.9, 19.5, and 17.0 may respectively. After evolving in Africa, Prodeinotherium likely migrated into Asia and then Europe with the formation of the "Gomphotherium land bridge". Prodeinotherium may have gone extinct around 15.5 mya, based on the last known fossils from the Arabian peninsula.[6] P. hobleyi was the first species to evolve, followed soon after P. pentapotamiae and then P. bavaricum around the same time. These species are from Africa, Europe and South Asia respectively.[2] Prodeinotherium was replaced in Asia by Deinotherium indicum, in Europe by D. giganteum, and in Africa by D. bozasi.[1][8]
Deinotheres are quite controversial with regards to the systematics. Many species have been named, yet major studies by Harris and Huttunen find that only three species in each genus are valid, based on distribution as well as smaller details. Within Prodeinotherium, the species found valid by these authors include P. bavaricum, P. pentapotamiae, and P. hobleyi. Many descriptions of new species of deinotheres are based upon limited material compared to only a small number of the species. Thus, many species of deinotheres are no longer valid.[1] In addition to invalid species, Prodeinotherium has on occasion been synonymized with Deinotherium. Species of Prodeinotherium found to be valid by multiple studies include P. cuvieri,[9] P. sinense,[10] and P. orlovii.[8] In a study 2011 analyzed the mandibles of multiple genera in a phylogenetic analysis. Their results are shown below:[11]
| Proboscidea |
| ||||||||||||||||||||||||||||||||||||||||||||||||
| |
History of discovery

Deinotherium bavaricum was originally mentioned in a paper by Christian Erich Hermann von Meyer in 1831. However, his first description of the material came in 1833, in which he also created the new species Deinotherium bavaricum, a lapsus calami. The material known is the lectotype P3, in the Bayerische Staatssammlung für Paläontologie und historische Geologie. From Bavaria, Meyer compared the tooth to the material of Deinotherium gigantium, and found enough features to distinguish it as a separate species. Most deinotheres were lumped into Deinotherium until the studies of Harris, who concluded that morphology separated them into two genera, Prodeinotherium and Deinotherium. As the earliest description of a small deinothere in Europe, P. bavaricum became the type species of Prodeinotherium.[1] Prodeinotherium was named in 1830 by Ehik, and its name is derived from pro – "before" Deinotherium – "terrible beast".[8]
Another early description of Prodeinotherium is that of Kaup (1832). He described teeth previously assigned to Tapir gigantesque, finding them to be a new deinothere. Kaup assigned these to Dinotherium cuvieri, using size to distinguish it from D. giganteum. However, the size, morphology, and distribution of P. bavaricum, and thus the latter became the senior synonym. Later in 1836, Lartet described yet another deinothere which eventually became P. bavaricum. This new species was named Deinotherium secondarium, for teeth from France. Lartet published no description, and did not mention this species in later works. Huttunen showed that the distribution of D. secondarium was within that of P. bavaricum, and thus considered the two species likely synonymous. Later, Ehik (1930) described the genus and the new species Prodinotherium hungaricum. The species was known from a jaw with teeth and some post cranial elements. This material was from Királd, and was destroyed, but casts of it remain in the Hungarian Natural History Museum. Diagnosed by dental features, and post cranial morphology, the specimen was later found similar to specimens from elsewhere in Europe, which were assigned to P. bavaricum. Harris found P. hungaricum to be a synonym of P. bavaricum, a conclusion followed by Huttunen. P. petenyii was described in 1989 by Vörös, who found it to differ from all other Prodeinotherium species. From Hungary, the material includes a jaw with teeth. The tooth morphology is very similar to that of P. bavaricum, and although the species has the unique feature of tusks that do not curve down and instead project forwards, Huttunen considered it a synonym of P. bavaricum.[1]
.jpg)
In 1868, Hugh Falconer's notes from before his death were published, including the description of material that he labelled Dinotherium pentapotamiae. This was from the Sewalik of India, he noted that the teeth were possibly the same as D. indium, but were within the range of a mid-sized individual. This presumption as D. indium was based on the possibility that D. giganteum was the only species of Deinotherium in Europe, and the variation of individuals of Mastodon longirostris within a population.[12] This species was later reassigned to Prodeinotherium, distinct from D. indicum.[1]
The African species, P. hobleyi, was first described in 1911 by Charles William Andrews. The species was from East Africa, and was named Dinotherium hobleyi. Andrews described these remains, which included a mandible with teeth, a calcaneum, a patella, and other indeterminable fragments, shipped to him by C.W. Hobley. It was compared to Dinotherium cuvieri, and although they were similar, the minor differences and geographical separation were enough for Andrews to create a new species.[13] This species was later reassigned to Prodeinotherium, as the only species from Africa.[1]
Prodeinotherium sinense was described in 2007 as a late species of Prodeinotherium it was described by Qui et al., and is known from dental material and jaw. It was first found in 2005, in Gansu, China. It is of a Late Miocene age, and is thus younger than other Prodeinotherium species. That study also found that the material of P. hungaricum is distinct from P. bavaricum.[14] A study in 2010 by Vergiev & Markow noted that the teeth are quite similar to those of Deinotherium, and based on these features and age the species was thought to either be a species in between Prodeinotherium and Deinotherium, or belonging to the latter genus.[10]
Early depictions of deinotheres like Prodeinotherium were scientifically incorrect. Before post cranial material was known, the genera were considered to be rhinos, giant tapirs, sirenians, whales and marsupials. However, with post cranial material came the proposal of an elephantine relation. However, early depictions of deinotheres were too elephantine, practically only with the addition of lower tusks. These restorations were inaccurate, because they showed the lower lip directly beneath the trunk, with the tusks projecting from the "chin". According to a 2001 study, it is more likely that the tusks projected above the lip, which followed the curvature of the jaw down. Another inaccuracy is likely the length of the trunks. Having a long elephantine trunk was thought of as unlikely by multiple authors, including Harris and the 2001 study. Besides the large opening often associated with a trunk, the general skull structure makes it unlikely for the trunk to be elongate. The upper tusks, retained in all more derived proboscideans, were likely lost so that the upper lip could directly manipulate the food of Prodeinotherium.[5]
Paleobiology

Prodeinotherium was a herbivorous organism. Based on the known distribution of fossils, Prodeinotherium could only survive along the coast in closed forests. Rodents and fish may have lived in the same environment or region as Prodeinotherium.[8] In Europe, fossils of Gomphotherium have been found alongside those of Prodeinotherium, showing that the genera likely ate different plants.[3]
Deinotheres were browsers, meaning they ate plants above ground level. It is possible that deinotheres specifically ate dicots. These could be found in closed woodland forests.[15][16] The way they chewed their food was probably similar to that of modern tapirs, with the front teeth being used to crush the food, while the second and third molars have a strong vertical shearing action, with little lateral (side to side) movement. This chewing action differs from both that of gomphotheres (lateral grinding) and elephants (horizontal shearing). Deinothere molars show little wear, indicating a diet of soft, nongritty, forest vegetation, with the down-turned lower tusks being used for stripping bark or other vegetation.[16] The supports for the tusks used in feeding is also based on the fact that juveniles have a different tusk morphology, which is consistent on them likely possessing a slightly different diet or feeding strategy. The trunk of deinotheres were likely similar to a tapirs, which could have been used for grasping plant matter and moving it to where the tongue could manipulate it.[5]
References
- 1 2 3 4 5 6 7 8 9 Huttunen, K. (2002). "Systematics and Taxonomy of the European Deinotheriidae (Proboscidea, Mammalia)" (PDF). Annalen des Naturhistorischen Museums in Wien. 103 A: 237–250.
- 1 2 3 4 5 Sanders, W. J., Kappelman, J. & Rasmussen, D. T., 2004 New large-bodied mammals from the late Oligocene site of Chilga, Ethiopia. Acta Palaeontologica Polonica Vol. 49, no.3, pp. 365–392
- 1 2 Huttunen, K.; Gohlich, U.B. (2002). "A partial skeleton of Prodeinotherium bavaricum (Proboscidea, Mammalia) from the Middle Miocene of Unterzolling (Upper Freshwater Molasse, Germany)" (PDF). Geobios 35: 489–514. doi:10.1016/s0016-6995(02)00042-6.
- 1 2 Larramendi, A. (2015). "Shoulder height, body mass and shape of proboscideans" (PDF). Acta Palaeontologica Polonica 60. doi:10.4202/app.00136.2014.
- 1 2 3 4 Markov, G.N.; Spassov, N.; Simeonovski, V. (2001). "A reconstruction of the facial morphology and feeding behaviour of the deinotheres". The World of Elephants – International Congress: 652–655.
- 1 2 3 Koufos, G.D.; Zouros, N.; Mourouzidou, O. (2003). "Prodeinotherium bavaricum (Proboscidea, Mammalia) from Lesvos island, Greece; the appearance of deinotheres in the Eastern Mediterranean" (PDF). Geobios 36: 305–315. doi:10.1016/S0016-6995(03)00031-7.
- ↑ Huttunen, K. (2004). "On a Prodeinotherium bavaricum (Proboscidea, Mammalia) skeleton from Franzensbad, Czech Republic" (PDF). Annalen des Naturhistorischen Museums in Wien. 105 A: 333–361.
- 1 2 3 4 Tiwari, B.N.; Verma, B.C.; Bhandari, A. (2006). "Record of Prodeinotherium (Proboscidea; Mammalia) from the Mid-Tertiary Dharmsala Group of the Kangra Valley, NW Himalaya, India: Biochronologic and Biogeographic Implications" (PDF). Journal of The Paleontological Society of India 51 (1): 93–100.
- ↑ Aiglstorfer, M.; Goehlich, U.B; Böhme, M.; Gross, M. (2014). "A partial skeleton of Deinotherium (Proboscidea, Mammalia) from the late Middle Miocene Gratkorn locality (Austria)" (PDF). Palaeobiodiversity and Palaeoenvironments 94 (2014). doi:10.1007/s12549-013-0140-x.
- 1 2 Vergiev, S.; Markov, G.N. (2010). "A mandible of Deinotherium (Mammalia: Proboscidea) from Aksakovo near Varna, Northeast Bulgaria" (PDF). Palaeodiversity 3: 241–247.
- ↑ Ferretti, M.P.; Debruyne, R. (2011). "Anatomy and phylogenetic value of the mandibular and coronoid canals and their associated foramina in proboscideans (Mammalia)" (PDF). Journal of the Linnaean Society 161: 391–413. doi:10.1111/j.1096-3642.2010.00637.x.
- ↑ Falconer, H.† (1868). Palaeontological Memoirs and Notes of the late Hugh Falconer: With a Biographical sketch of the Author 2. Sottiswoode & Co. p. 5.
- ↑ Andrews, C.W. (1911). "On a new species of Dinotherium (Dinotherium hobleyi) from British East Africa". Proceedings of The Zoological Society of London: 943–945. doi:10.1111/j.1096-3642.1911.tb01964.x.
- ↑ Qiu, Z.X.; Wang, B.W.; Li, H.; Deng, T.; Sun, Y. (2007). "First discovery of a deinothere in China". Vertebrata PalAsiatica 45 (4): 261–277.
- ↑ Calandra, I.; Göhlich, U.B.; Merceron, G. (2010). "Feeding preferences of Gomphotherium subtapiroideum (Proboscidea, Mammalia) from the Miocene of Sandelzhausen (Northern Alpine Foreland Basin, southern Germany) through life and geological time: evidence from dental microwear analysis". Paläontologische Zeitschrift 84 (2010): 205–215. doi:10.1007/s12542-010-0054-0.
- 1 2 Harris, J.M. (1975). "Evolution of feeding mechanisms in the family Deinotheriidae (Mammalia: Proboscidea)". Zoological Journal of the Linnaean Society (56): 331–362.
External links
- Phenomena: The largest Beast to walk the Earth discussing the size and weight estimates of Larramendi (2015).