Pleckstrin homology domain
|
PH domain of tyrosine-protein kinase BTK | |||||||||
| Identifiers | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Symbol | PH | ||||||||
| Pfam | PF00169 | ||||||||
| InterPro | IPR001849 | ||||||||
| SMART | PH | ||||||||
| PROSITE | PDOC50003 | ||||||||
| SCOP | 1dyn | ||||||||
| SUPERFAMILY | 1dyn | ||||||||
| OPM superfamily | 51 | ||||||||
| OPM protein | 1pls | ||||||||
| CDD | cd00821 | ||||||||
| |||||||||
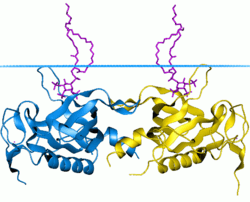
Pleckstrin homology domain (PH domain) is a protein domain of approximately 120 amino acids that occurs in a wide range of proteins involved in intracellular signaling or as constituents of the cytoskeleton.[1][2][3][4][5][6][7]
This domain can bind Phosphatidylinositol lipids within biological membranes (such as Phosphatidylinositol (3,4,5)-trisphosphate and phosphatidylinositol (4,5)-bisphosphate),[8] and proteins such as the βγ-subunits of heterotrimeric G proteins,[9] and protein kinase C.[10] Through these interactions, PH domains play a role in recruiting proteins to different membranes, thus targeting them to appropriate cellular compartments or enabling them to interact with other components of the signal transduction pathways.
Lipid binding specificity
Individual PH domains possess specificities for phosphoinositides phosphorylated at different sites within the inositol ring, e.g., some bind phosphatidylinositol (4,5)-bisphosphate but not phosphatidylinositol (3,4,5)-trisphosphate or phosphatidylinositol (3,4)-bisphosphate, while others may possess the requisite affinity. This is important because it makes the recruitment of different PH domain containing proteins sensitive to the activities of enzymes that either phosphorylate or dephosphorylate these sites on the inositol ring, such as phosphoinositide 3-kinase or PTEN, respectively. Thus, such enzymes exert a part of their effect on cell function by modulating the localization of downstream signaling proteins that possess PH domains that are capable of binding their phospholipid products.
Structure
The 3D structure of several PH domains has been determined.[11] All known cases have a common structure consisting of two perpendicular anti-parallel beta sheets, followed by a C-terminal amphipathic helix. The loops connecting the beta-strands differ greatly in length, making the PH domain relatively difficult to detect while providing the source of the domain's specificity. The only conserved residue among PH domains is a single tryptophan located within the alpha helix that serves to nucleate the core of the domain.
Proteins containing PH domain
PH domains can be found in many different proteins, such as OSBP or ARF. Recruitment to the Golgi in this case is dependent on both PtdIns and ARF. A large number of PH domains have poor affinity for phosphoinositides and are hypothesized to function as protein binding domains. A Genome-wide look in Saccharomyces cerevisiae showed that most of the 33 yeast PH domains are indeed promiscuous in binding to phosphoinositides, while only one (Num1-PH) behaved highly specific .[12] Proteins reported to contain PH domains belong to the following families:
- Pleckstrin, the protein where this domain was first detected, is the major substrate of protein kinase C in platelets. Pleckstrin is one of the rare proteins to contain two PH domains.
- Ser/Thr protein kinases such as the Akt/Rac family, the beta-adrenergic receptor kinases, the mu isoform of PKC and the trypanosomal NrkA family.
- Tyrosine protein kinases belonging to the Btk/Itk/Tec subfamily.
- Insulin Receptor Substrate 1 (IRS-1).
- Regulators of small G-proteins like guanine nucleotide releasing factor GNRP (Ras-GRF) (which contains 2 PH domains), guanine nucleotide exchange proteins like vav, dbl, SoS and S. cerevisiae CDC24, GTPase activating proteins like rasGAP and BEM2/IPL2, and the human break point cluster protein bcr.
- Cytoskeletal proteins such as dynamin (see IPR001401), Caenorhabditis elegans kinesin-like protein unc-104 (see IPR001752), spectrin beta-chain, syntrophin (2 PH domains), and S. cerevisiae nuclear migration protein NUM1.
- Mammalian phosphatidylinositol-specific phospholipase C (PI-PLC) (see IPR000909) isoforms gamma and delta. Isoform gamma contains two PH domains, the second one is split into two parts separated by about 400 residues.
- Oxysterol-binding proteins OSBP, S. cerevisiae OSH1 and YHR073w.
- Mouse protein citron, a putative rho/rac effector that binds to the GTP-bound forms of rho and rac.
- Several S. cerevisiae proteins involved in cell cycle regulation and bud formation like BEM2, BEM3, BUD4 and the BEM1-binding proteins BOI2 (BEB1) and BOI1 (BOB1).
- C. elegans protein MIG-10.
- Ceramide kinase, a lipid kinase that phosphorylates ceramides to ceramide-1-phosphate.[13]
Subfamilies
Examples
Human genes encoding proteins containing this domain include:
- ABR, ADRBK1, ADRBK2, AFAP, AFAP1, AFAP1L1, AFAP1L2, AKAP13, AKT1, AKT2, AKT3, ANLN, APBB1IP, APPL1, APPL2, ARHGAP10, ARHGAP12, ARHGAP15, ARHGAP21, ARHGAP22, ARHGAP23, ARHGAP24, ARHGAP25, ARHGAP26, ARHGAP27, ARHGAP9, ARHGEF16, ARHGEF18, ARHGEF19, ARHGEF2, ARHGEF3, ARHGEF4, ARHGEF5, ARHGEF6, ARHGEF7, ARHGEF9, ASEF2,
- BMX, BTK,
- C20orf42, C9orf100, CADPS, CADPS2, CDC42BPA, CDC42BPB, CDC42BPG, CENTA1, CENTA2, CENTB1, CENTB2, CENTB5, CENTD1, CENTD2, CENTD3, CENTG1, CENTG2, CENTG3, CIT, CNKSR1, CNKSR2, COL4A3BP, CTGLF1, CTGLF2, CTGLF3, * CTGLF4, CTGLF5, CTGLF6,
- DAB2IP, DAPP1, DDEF1, DDEF2, DDEFL1, DEF6, DEPDC2, DGKD, DGKH, DGKK, DNM1, DNM2, DNM3, DOCK10, DOCK11, DOCK9, DOK1, DOK2, DOK3, DOK4, DOK5, DOK6, DTGCU2,
- EXOC8,
- FAM109A, FAM109B, FARP1, FARP2, FGD1, FGD2, FGD3, FGD4, FGD5, FGD6,
- GAB1, GAB2, GAB3, GAB4, GRB10, GRB14, GRB7,
- IRS1, IRS2, IRS4, ITK, ITSN1, ITSN2,
- KALRN, KIF1A, KIF1B, KIF1Bbeta,
- MCF2, MCF2L, MCF2L2, MRIP, MYO10,
- NET1, NGEF,
- OBPH1, OBSCN, OPHN1, OSBP, OSBP2, OSBPL10, OSBPL11, OSBPL3, OSBPL5, OSBPL6, OSBPL7, OSBPL8, OSBPL9,
- PHLDA2, PHLDA3, PHLDB1, PHLDB2, PHLPP, PIP3-E, PLCD1, PLCD4, PLCG1, PLCG2, PLCH1, PLCH2, PLCL1, PLCL2, PLD1, PLD2, PLEK, PLEK2, PLEKHA1, PLEKHA2, PLEKHA3, PLEKHA4, PLEKHA5, PLEKHA6, PLEKHA7, PLEKHA8, PLEKHB1, PLEKHB2, PLEKHC1, PLEKHF1, PLEKHF2, PLEKHG1, PLEKHG2, PLEKHG3, PLEKHG4, PLEKHG5, PLEKHG6, PLEKHH1, PLEKHH2, PLEKHH3, PLEKHJ1, PLEKHK1, PLEKHM1, PLEKHM2, PLEKHO1, PLEKHQ1, PREX1, PRKCN, PRKD1, PRKD2, PRKD3, PSCD1, PSCD2, PSCD3, PSCD4, PSD, PSD2, PSD3, PSD4, RALGPS1, RALGPS2, RAPH1,
- RASA1, RASA2, RASA3, RASA4, RASAL1, RASGRF1, RGNEF, ROCK1, ROCK2, RTKN,
- SBF1, SBF2, SCAP2, SGEF, SH2B, SH2B1, SH2B2, SH2B3, SH3BP2, SKAP1, SKAP2, SNTA1, SNTB1, SNTB2, SOS1, SOS2, SPATA13, SPNB4, SPTBN1, SPTBN2, SPTBN4, SPTBN5, STAP1, SWAP70, SYNGAP1,
- TBC1D2, TEC, TIAM1, TRIO, TRIOBP, TYL,
- URP1, URP2,
- VAV1, VAV2, VAV3, VEPH1
See also
- Pleckstrin
- The unrelated FYVE domain binds Phosphatidylinositol 3-phosphate and has been found in over 60 proteins.
- The GRAM domain is a structurally related protein domain.
References
- ↑ Mayer, B. J.; Ren, R.; Clark, K. L.; Baltimore, D. (1993). "A putative modular domain present in diverse signaling proteins". Cell 73 (4): 629–630. doi:10.1016/0092-8674(93)90244-K. PMID 8500161.
- ↑ Haslam, R. J.; Koide, H. B.; Hemmings, B. A. (1993). "Pleckstrin domain homology". Nature 363 (6427): 309–10. doi:10.1038/363309b0. PMID 8497315.
- ↑ Musacchio, A.; Gibson, T.; Rice, P.; Thompson, J.; Saraste, M. (1993). "The PH domain: A common piece in the structural pathcwork of signalling proteins". Trends in Biochemical Sciences 18 (9): 343–348. doi:10.1016/0968-0004(93)90071-T. PMID 8236453.
- ↑ Gibson, T. J.; Hyvönen, M.; Musacchio, A.; Saraste, M.; Birney, E. (1994). "PH domain: The first anniversary". Trends in Biochemical Sciences 19 (9): 349–353. doi:10.1016/0968-0004(94)90108-2. PMID 7985225.
- ↑ Pawson, T. (1995). "Protein modules and signalling networks". Nature 373 (6515): 573–580. doi:10.1038/373573a0. PMID 7531822.
- ↑ Ingley, E.; Hemmings, B. A. (1994). "Pleckstrin homology (PH) domains in signal transducton". Journal of Cellular Biochemistry 56 (4): 436–443. doi:10.1002/jcb.240560403. PMID 7890802.
- ↑ Saraste, M.; Hyvönen, M. (1995). "Pleckstrin homology domains: A fact file". Current Opinion in Structural Biology 5 (3): 403–408. doi:10.1016/0959-440X(95)80104-9. PMID 7583640.
- ↑ Wang, D. S.; Shaw, G. (1995). "The Association of the C-Terminal Region of β1ΣII Spectrin to Brain Membranes is Mediated by a pH Domain, Does Not Require Membrane Proteins, and Coincides with a Inositol-1,4,5 Trisphosphate Binding Site". Biochemical and Biophysical Research Communications 217 (2): 608–615. doi:10.1006/bbrc.1995.2818. PMID 7503742.
- ↑ Wang, D. S.; Shaw, R.; Winkelmann, J. C.; Shaw, G. (1994). "Binding of PH Domains of β-Adrenergic-Receptor Kinase and β-Spectrin to WD40/β-Transducin Repeat Containing Regions of the β-Subunit of Trimeric G-Proteins". Biochemical and Biophysical Research Communications 203 (1): 29–35. doi:10.1006/bbrc.1994.2144. PMID 8074669.
- ↑ Yao, L.; Kawakami, Y.; Kawakami, T. (1994). "The pleckstrin homology domain of Bruton tyrosine kinase interacts with protein kinase C". Proceedings of the National Academy of Sciences 91 (19): 9175–9179. doi:10.1073/pnas.91.19.9175.
- ↑ Riddihough, G. (1994). "More meanders and sandwiches". Nature Structural Biology 1 (11): 755–757. doi:10.1038/nsb1194-755. PMID 7634082.
- ↑ Yu, J. W.; Mendrola, J. M.; Audhya, A.; Singh, S.; Keleti, D.; Dewald, D. B.; Murray, D.; Emr, S. D.; Lemmon, M. A. (2004). "Genome-Wide Analysis of Membrane Targeting by S. Cerevisiae Pleckstrin Homology Domains". Molecular Cell 13 (5): 677–688. doi:10.1016/S1097-2765(04)00083-8. PMID 15023338.
- ↑ Sugiura, M.; Kono, K.; Liu, H.; Shimizugawa, T.; Minekura, H.; Spiegel, S.; Kohama, T. (2002). "Ceramide Kinase, a Novel Lipid Kinase. MOLECULAR CLONING AND FUNCTIONAL CHARACTERIZATION". Journal of Biological Chemistry 277 (26): 23294–23300. doi:10.1074/jbc.M201535200. PMID 11956206.
External links
- Nash Lab Protein Interaction Domains - PH domain description
- UMich Orientation of Proteins in Membranes families/superfamily-51 - Calculated orientations of PH domains in membranes