Myxococcus xanthus
| Myxococcus xanthus | |
|---|---|
 | |
| Scientific classification | |
| Kingdom: | Bacteria |
| Phylum: | Proteobacteria |
| Class: | Delta Proteobacteria |
| Order: | Myxococcales |
| Family: | Myxococcaceae |
| Genus: | Myxococcus |
| Species: | M. xanthus |
| Binomial name | |
| Myxococcus xanthus Beebe 1941 | |
Myxococcus xanthus is a gram-negative, rod-shaped species of myxobacteria that exhibits various forms of self-organizing behavior as a response to environmental cues. Under normal conditions with abundant food, it exists as a predatory, saprophytic single-species biofilm called a swarm. Under starvation conditions, it undergoes a multicellular development cycle.[1]
Colony growth
A swarm of M. xanthus is a distributed system, containing millions of entities that communicate among themselves in a non-centralized fashion. Simple patterns of cooperative behavior among the members of the colony combine to generate complex group behaviors in a process known as "stigmergy". For example, the tendency for one cell to glide only when in direct contact with another results in the colony forming swarms called "wolf-packs" that may up to several inches wide. This behavior is advantageous to the members of the swarm, as it increases the concentration of extracellular digestive enzymes secreted by the bacteria, thus facilitating predatory feeding. Likewise, during stressful conditions, the bacteria undergo a process in which about 100,000 individual cells aggregate to form a structure called the fruiting body over the course of several hours. On the interior of the fruiting body, the rod-shaped cells differentiate into spherical, thick-walled spores. They undergo changes in the synthesis of new proteins, as well as alterations in the cell wall, which parallel the morphological changes. During these aggregations, dense ridges of cells move in ripples, which wax and wane over 5 hours.[2]
Motility
An important part of M. xanthus behavior is its ability to move on a solid surface by a mechanism called "gliding".[3] Gliding Motility is a method of locomotion that allows for movement, without the help of a flagella, on a solid surface. Gliding Motility is separated into two groups for the M. xanthus: A-motility (adventurous) and S-motility (social).[4] In A motility, single cells move, resulting in a distribution with many single cells. In S motility, single cells do not move, but cells that are close to one another move. This leads to a spatial distribution of cells with lots of clusters and few isolated single cells.[4]
More than 37 genes are involved in the A-motility system, which comprises multiple motor elements that are arrayed along the entire cell body. Each motor element appears to be localized to the periplasmic space and is bound to the peptidoglycan layer. The motors are hypothesized to move on helical cytoskeletal filaments. Gliding force generated by these motors is coupled to adhesion sites that move freely in the outer membrane, and which provide a specific contact with the substratum, possibly aided by extracellular polysaccharide slime.[5][6]
S-motility may represent a variation of twitching motility, since it is mediated by the extension and retraction of type IV pili that extend through the leading cell pole. The genes of the S-motility system appear to be homologs of genes involved in the biosynthesis, assembly, and function of twitching motility in other bacteria.[5][6]
Cell differentiation, fruiting and sporulation
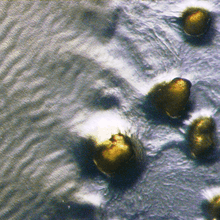
In response to starvation, myxobacteria develop species-specific multicellular fruiting bodies. Starting from a uniform swarm of cells, some aggregate into fruiting bodies, while other cells remain in a vegetative state. Those cells that participate in formation of the fruiting body transform from rods into spherical, heat-resistant myxospores, while the peripheral cells remain rod-shaped.[7] Although not as tolerant to environmental extremes as, say, Bacillus endospores, the relative resistance of myxospores to desiccation and freezing enables myxobacteria to survive seasonally harsh environments. When a nutrient source becomes once again available, the myxospores germinate, shedding their spore coats to emerge into rod-shaped vegetative cells. The synchronized germination of thousands of myxospores from a single fruiting body enables the members of the new colony of myxobacteria to immediately engage in cooperative feeding.[8]
Intercellular communication
It is very likely that cells communicate during the process of fruiting and sporulation, because a group of cells that starved together form myxospores inside fruiting bodies.[1] Intercellular signal appears to be necessary to ensure that sporulation happens in the proper place and at the proper time.[9] Research supports the existence of an extracellular signal, A-factor, which is necessary for developmental gene expression and for the development of a complete fruiting body.[10]
Importance in research
The complex life cycles of the myxobacteria make them very attractive models for the study of gene regulation as well as cell to cell interactions. The traits of M. xanthus make it very easy to study, and therefore important to research. Laboratory strains of M. xanthus are available that are capable of planktonic growth in shaker culture, so that they are easy to grow in large numbers. The tools of classical and molecular genetics are relatively well-developed in M. xanthus.[11]
Although the fruiting bodies of M. xanthus are relatively primitive compared with, say, the elaborate structures produced by Stigmatella aurantiaca and other myxobacteria, the great majority of genes known to be involved in development are conserved across species.[12] In order to make agar cultures of M. xanthus grow into fruiting bodies, one simply can plate the bacteria on starvation media.[13] Furthermore, it is possible to artificially induce the production of myxospores without the intervening formation of fruiting bodies, by adding compounds such as glycerol or various metabolites to the medium.[14] In this way, different stages in the developmental cycle can be experimentally isolated.
The genome of M. xanthus has been completely sequenced.[15] The size of its genome may reflect the complexity of its life cycle. At 9.14 megabase, it had the largest known prokaryotic genome until the sequencing of Sorangium cellulosum (12.3 Mb), which is also a myxobacterium.
Developmental cheating
Social cheating exists among M. Xanthus commonly. As long as mutants are not too common, if they are unable to perform the group beneficial function of producing spores, they will still reap the benefit of the population as a whole. Research has shown that 4 different types of M. Xanthus mutants showed forms of cheating during development, by being over-represented among spores relative to their initial frequency in the mixture.[16]
M. xanthus and evolution
In 2003, two scientists, Velicer and Yu, deleted certain parts of the M. xanthus genome, making it unable to swarm effectively on soft agar. Individuals were cloned, and allowed to evolve. After a period of 64 weeks, two of the evolving populations had started to swarm outward almost as effectively as normal wild-type colonies. However, the patterns of the swarm were very different from those of the wild-type bacteria. This suggested that they had developed a new way of moving, and Velicer and Yu confirmed this by showing that the new populations had not regained the ability to make pili, which allows wild-type bacteria to swarm. This study addressed questions about the evolution of cooperation between individuals that had plagued scientists for years.
Strains
- Myxococcus xanthus DK 1622
- Myxococcus xanthus DZ2
- Myxococcus xanthus DZF1
- Myxococcus xanthus NewJersey2
References
- 1 2 Kroos, Lee; Kuspa, Adam; Kaiser, Dale (1986). "A global analysis of developmentally regulated genes in Myxococcus xanthus". Developmental Biology 117 (1): 252–266. doi:10.1016/0012-1606(86)90368-4. PMID 3017794. Retrieved 26 May 2015.
- ↑ Velicer GJ, Stredwick KL (2002). "Experimental social evolution with Myxococcus xanthus". Antonie Van Leeuwenhoek 81 (1–4): 155–64. doi:10.1023/A:1020546130033. PMID 12448714.
- ↑ Shi, Wenyuan. "Myxococcus xanthus". University of California at Los Angeles. Retrieved 26 May 2015.
- 1 2 Kaiser, Dale (1979). "Social gliding is correlated with the presence of pili in Myxococcus xanthus" (PDF). Proc. Nat. Acad. Sci. 76 (11): 5952–5956. doi:10.1073/pnas.76.11.5952. PMC 411771. PMID 42906. Retrieved 26 May 2015.
- 1 2 Spormann, A. M. (1999). "Gliding motility in bacteria: insights from studies of Myxococcus xanthus". Microbiol. Mol. Biol. Rev. 63 (3): 621–641. PMC 103748. PMID 10477310.
- 1 2 Zusman, D. R.; Scott, A. E.; Yang, Z.; Kirby, J. R. (2007). "Chemosensory pathways, motility and development in Myxococcus xanthus". Nature Reviews Microbiology 5 (11): 862–72. doi:10.1038/nrmicro1770. PMID 17922045.
- ↑ Julien, B.; Kaiser, A. D.; Garza, A. (2000). "Spatial control of cell differentiation in Myxococcus xanthus". Proceedings of the National Academy of Sciences 97 (16): 9098–9103. doi:10.1073/pnas.97.16.9098.
- ↑ Lee, Keesoo; Shimkets, Lawrence J. (1994). "Cloning and Characterization of the socA Locus Which Restores Development to Myxococcus xanthus C-Signaling Mutants" (PDF). Journal of Bacteriology 176 (8): 2200–2209. PMID 8157590. Retrieved 30 May 2015.
- ↑ Hagan, David C.; Bretscher, Anthony P.; Kaiser, Dale (1978). "Synergism between morphogenetic mutants of Myxococcus xanthus". Developmental Biology 64 (2): 284–296. doi:10.1016/0012-1606(78)90079-9. PMID 98366. Retrieved 26 May 2015.
- ↑ Kuspa, Adam; Kroos, Lee; Kaiser, Dale (1986). "Intercellular signaling is required for developmental gene expression in Myxococcus xanthus". Developmental Biology 117 (1): 267–276. doi:10.1016/0012-1606(86)90369-6. PMID 3017795. Retrieved 26 May 2015.
- ↑ Stephens, Karen; Kaiser, Dale (1987). "Genetics of gliding motility in Myxococcus xanthus: Molecular cloning of the mgl locus". Molecular and General Genetics 207: 256–266. doi:10.1007/bf00331587.
- ↑ Huntley, S.; Hamann, N.; Wegener-Feldbrugge, S.; Treuner-Lange, A.; Kube, M.; Reinhardt, R.; Klages, S.; Muller, R.; Ronning, C. M.; Nierman, W. C.; Sogaard-Andersen, L. (2010). "Comparative Genomic Analysis of Fruiting Body Formation in Myxococcales". Molecular Biology and Evolution 28 (2): 1083–1097. doi:10.1093/molbev/msq292. PMID 21037205.
- ↑ Inouye, Masayori; Inouye, Sumiko; Zusman, David R. (1979). "Gene expression during development of Myxococcus xanthus: Pattern of protein synthesis". Developmental Biology 68 (2): 579–591. doi:10.1016/0012-1606(79)90228-8. PMID 108160. Retrieved 26 May 2015.
- ↑ Bui, N. K.; Gray, J.; Schwarz, H.; Schumann, P.; Blanot, D.; Vollmer, W. (2008). "The Peptidoglycan Sacculus of Myxococcus xanthus Has Unusual Structural Features and is Degraded during Glycerol-Induced Myxospore Development". Journal of Bacteriology 191 (2): 494–505. doi:10.1128/JB.00608-08. PMID 18996994.
- ↑ Goldman, B. S.; Nierman, W. C.; Kaiser, D.; Slater, S. C.; Durkin, A. S.; Eisen, J. A.; Ronning, C. M.; Barbazuk, W. B.; Blanchard, M.; Field, C.; Halling, C.; Hinkle, G.; Iartchuk, O.; Kim, H. S.; MacKenzie, C.; Madupu, R.; Miller, N.; Shvartsbeyn, A.; Sullivan, S. A.; Vaudin, M.; Wiegand, R.; Kaplan, H. B. (2006). "Evolution of sensory complexity recorded in a myxobacterial genome". Proceedings of the National Academy of Sciences 103 (41): 15200–15205. doi:10.1073/pnas.0607335103.
- ↑ Velicer, G. J.; Kroos, L.; Lenski, R. E. (2000). "Developmental cheating in the social bacterium Myxococcus xanthus". Nature 404 (6778): 598–601. doi:10.1038/35007066. PMID 10766241.
External links
- Model Organism Database
- John Kirby at the University of Iowa
- Dale Kaiser Lab at Stanford University
- Watching social behaviour evolve
- Taxonomic Information for Myxococcus xanthus
Videos
- Myxococcus xanthus preying on an E. coli colony
- Myxococcus xanthus fruiting body formation
- Myxococcus xanthus ripples – Predation
- Predatory bacterial crowdsourcing