Mutation–selection balance
Mutation–selection balance is an equilibrium in the number of deleterious alleles in a population that occurs when the rate at which deleterious alleles are created by mutation equals the rate at which deleterious alleles are eliminated by selection.[1][2][3][4] The majority of genetic mutations are neutral or deleterious; beneficial mutations are relatively rare. The resulting influx of deleterious mutations into a population over time is counteracted by negative selection, which acts to purge deleterious mutations. Setting aside other factors (e.g. balancing selection and genetic drift), the equilibrium number of deleterious alleles is then determined by a balance between the deleterious mutation rate and the rate at which selection purges those mutations.
Mutation–selection balance was originally proposed to explain how genetic variation is maintained in populations, although several other ways for deleterious mutations to persist are now recognized, notably balancing selection.[5] Nevertheless, the concept is still widely used in evolutionary genetics e.g. to explain the persistence of deleterious alleles in actual populations as in the case of spinal muscular atrophy,[6][7] or, in theoretical models, mutation-selection balance can appear in a variety of ways and has even been applied to beneficial mutations (i.e. balance between selective loss of variation and creation of variation by beneficial mutations).[8]
Haploid population
As a simple example of mutation-selection balance, consider a single locus in a haploid population with two possible alleles: a normal allele A with frequency 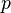 , and a mutated deleterious allele B with frequency
, and a mutated deleterious allele B with frequency 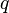 , which has a small relative fitness disadvantage of
, which has a small relative fitness disadvantage of  . Suppose that deleterious mutations from A to B occur at rate
. Suppose that deleterious mutations from A to B occur at rate 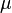 , and the reverse beneficial mutation from B to A occurs rarely enough to be negligible (e.g. because the mutation rate is so low that is small). Then, each generation selection eliminates deleterious mutants thereby reducing by an amount
, and the reverse beneficial mutation from B to A occurs rarely enough to be negligible (e.g. because the mutation rate is so low that is small). Then, each generation selection eliminates deleterious mutants thereby reducing by an amount 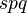 , while mutation creates more deleterious alleles thereby increasing by an amount
, while mutation creates more deleterious alleles thereby increasing by an amount 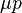 . Mutation–selection balance occurs when these forces cancel and is constant from generation to generation, implying
. Mutation–selection balance occurs when these forces cancel and is constant from generation to generation, implying 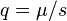 .[9] Thus, provided that the mutant allele is not weakly deleterious (very small ) and the mutation rate is not very high, the equilibrium frequency of the deleterious allele will be small.
.[9] Thus, provided that the mutant allele is not weakly deleterious (very small ) and the mutation rate is not very high, the equilibrium frequency of the deleterious allele will be small.
Diploid population with random mating
In a diploid population, a deleterious allele B may have different effects on individual fitness in heterozygotes AB and homozygotes BB depending on the degree of dominance of the normal allele A. To represent this, let the relative fitness of deleterious homozygotes and heterozygotes be smaller than that of normal homozygotes AA by factors of 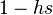 and
and 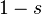 respectively, where
respectively, where  is a number between
is a number between  and
and 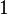 measuring the degree of dominance (
measuring the degree of dominance (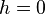 indicates that A is completely dominant while
indicates that A is completely dominant while 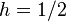 indicates no dominance).
indicates no dominance).
The degree of dominance affects the relative importance of selection on heterozygotes versus homozygotes. If A is not completely dominant (i.e. is not close to zero), then deleterious mutations are primarily removed by selection on heterozygotes because they contain the vast majority of deleterious B alleles (provided that the deleterious mutation rate is much smaller than ). This case is approximately equivalent to the preceding haploid case, where mutation converts normal homozygotes to heterozygotes at rate and selection acts on heterzygotes with selection coefficient 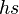 ; thus
; thus 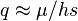 .[10]
.[10]
In the case of complete dominance (), deleterious alleles are only removed by selection on BB homozygotes. Let 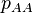 ,
, 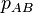 and
and 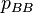 be the frequencies of the corresponding genotypes. The frequency
be the frequencies of the corresponding genotypes. The frequency 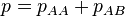 of normal alleles A increases at rate
of normal alleles A increases at rate 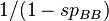 due to the selective elimination of recessive homozygotes, while mutation causes to shrink at rate
due to the selective elimination of recessive homozygotes, while mutation causes to shrink at rate 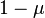 (again we ignore back mutations). Mutation–selection balance occurs when these rates balance and
(again we ignore back mutations). Mutation–selection balance occurs when these rates balance and 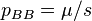 . Since mating is random, this implies that the frequency of deleterious alleles is
. Since mating is random, this implies that the frequency of deleterious alleles is 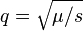 .[11] This equilibrium frequency is potentially substantially larger than for the preceding case of partial dominance, reflecting the reservoir of mutant alleles carried in unaffected heterozygotes which are shielded from selection.
.[11] This equilibrium frequency is potentially substantially larger than for the preceding case of partial dominance, reflecting the reservoir of mutant alleles carried in unaffected heterozygotes which are shielded from selection.
See also
References
- ↑ Kimura, James F. Crow, Motoo (1970). An introduction to population genetics theory ([Reprint] ed.). New Jersey: Blackburn Press. ISBN 9781932846126.
- ↑ Lynch, Michael (August 2010). "Evolution of the mutation rate". Trends in Genetics 26 (8): 345–352. doi:10.1016/j.tig.2010.05.003. PMC 2910838. PMID 20594608.
- ↑ Barton, Nicholas H. (2007). Evolution. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press. ISBN 9780879696849.
- ↑ Herron, JC and S Freeman. 2014. Evolutionary Analysis, 5th Edition. Pearson.
- ↑ Barton, Nicholas H. (2007). Evolution. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press. ISBN 9780879696849.
- ↑ "De Novo Rearrangements Found in 2% of Index Patients with Spinal Muscular Atrophy: Mutational Mechanisms, Parental Origin, Mutation Rate, and Implications for Genetic Counseling". The American Journal of Human Genetics 61: 1102–1111. doi:10.1086/301608.
- ↑ Herron, JC and S Freeman. 2014. Evolutionary Analysis, 5th Edition. Pearson.
- ↑ http://www.genetics.org/content/176/3/1759.full
- ↑ Barton, Nicholas H. (2007). Evolution. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press. ISBN 9780879696849.
- ↑ Kimura, James F. Crow, Motoo (1970). An introduction to population genetics theory ([Reprint] ed.). New Jersey: Blackburn Press. ISBN 9781932846126.
- ↑ Kimura, James F. Crow, Motoo (1970). An introduction to population genetics theory ([Reprint] ed.). New Jersey: Blackburn Press. ISBN 9781932846126.