Leslie matrix
In applied mathematics, the Leslie matrix is a discrete, age-structured model of population growth that is very popular in population ecology. It was invented by and named after Patrick H. Leslie. The Leslie matrix (also called the Leslie model) is one of the most well known ways to describe the growth of populations (and their projected age distribution), in which a population is closed to migration, growing in an unlimited environment, and where only one sex, usually the female, is considered.
The Leslie matrix is used in ecology to model the changes in a population of organisms over a period of time. In a Leslie model, the population is divided into groups based on age classes. A similar model which replaces age classes with ontogenetic stages is called a Lefkovitch matrix,[1] whereby individuals can both remain in the same stage class or move on to the next one. At each time step, the population is represented by a vector with an element for each age class where each element indicates the number of individuals currently in that class.
The Leslie matrix is a square matrix with the same number of rows and columns as the population vector has elements. The (i,j)th cell in the matrix indicates how many individuals will be in the age class i at the next time step for each individual in stage j. At each time step, the population vector is multiplied by the Leslie matrix to generate the population vector for the subsequent time step.
To build a matrix, some information must be known from the population:
-
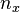 , the count of individuals (n) of each age class x
, the count of individuals (n) of each age class x -
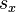 , the fraction of individuals that survives from age class x to age class x+1,
, the fraction of individuals that survives from age class x to age class x+1, -
 , fecundity, the per capita average number of female offspring reaching
, fecundity, the per capita average number of female offspring reaching 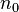 born from mother of the age class x. More precisely, it can be viewed as the number of offspring produced at the next age class
born from mother of the age class x. More precisely, it can be viewed as the number of offspring produced at the next age class 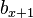 weighted by the probability of reaching the next age class. Therefore
weighted by the probability of reaching the next age class. Therefore 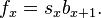
From the observations that at time t+1 is simply the sum of all offspring born from the previous time step and that the organisms surviving to time t+1 are the organisms at time t surviving at probability , one gets 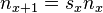 . This then implies the following matrix representation:
. This then implies the following matrix representation:
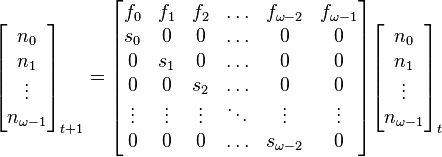
where 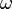 is the maximum age attainable in the population.
is the maximum age attainable in the population.
This can be written as:
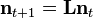
or:
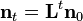
where 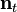 is the population vector at time t and
is the population vector at time t and 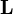 is the Leslie matrix. The dominant eigenvalue of , denoted
is the Leslie matrix. The dominant eigenvalue of , denoted  , gives the population's asymptotic growth rate (growth rate at the stable age distribution). The corresponding eigenvector provides the stable age distribution, the proportion of individuals of each age within the population. Once the stable age distribution has been reached, a population undergoes exponential growth at rate .
, gives the population's asymptotic growth rate (growth rate at the stable age distribution). The corresponding eigenvector provides the stable age distribution, the proportion of individuals of each age within the population. Once the stable age distribution has been reached, a population undergoes exponential growth at rate .
The characteristic polynomial of the matrix is given by the Euler–Lotka equation.
The Leslie model is very similar to a discrete-time Markov chain. The main difference
is that in a Markov model, one would have 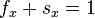 for each
for each  ,
while the Leslie model may have these sums greater or less than 1.
,
while the Leslie model may have these sums greater or less than 1.
Stable age structure
This age-structured growth model suggests a steady-state, or stable, age-structure and growth rate. Regardless of the initial population size,  , or age distribution, the population tends asymptotically to this age-structure and growth rate. It also returns to this state following perturbation. The Euler–Lotka equation provides a means of identifying the intrinsic growth rate. The stable age-structure is determined both by the growth rate and the survival function (i.e. the Leslie matrix). For example, a population with a large intrinsic growth rate will have a disproportionately “young” age-structure. A population with high mortality rates at all ages (i.e. low survival) will have a similar age-structure. Charlesworth (1980) provides further details on the rate and form of convergence to the stable age-structure.
, or age distribution, the population tends asymptotically to this age-structure and growth rate. It also returns to this state following perturbation. The Euler–Lotka equation provides a means of identifying the intrinsic growth rate. The stable age-structure is determined both by the growth rate and the survival function (i.e. the Leslie matrix). For example, a population with a large intrinsic growth rate will have a disproportionately “young” age-structure. A population with high mortality rates at all ages (i.e. low survival) will have a similar age-structure. Charlesworth (1980) provides further details on the rate and form of convergence to the stable age-structure.
The random Leslie case
There is a generalization of the population growth rate to when a Leslie matrix has random elements may be correlated. When characterizing the disorder, or uncertainties, in vital parameters; a perturbative formalism to deal with linear non-negative random matrix difference equations has to be used. Then: the non-trivial, effective eigenvalue which defines the long-time asymptotic dynamics of the mean-value population vector state can be presented as the effective growth rate. This eigenvalue and the associated mean-value invariant vector state can be calculated from the smallest positive root of a secular polynomial and the residue of the mean-value Green function. Exact and perturbative results can be analyzed for several models of disorder.
References
- Caceres, M.O. and Caceres-Saez, I. 2011. Random Leslie Matrices in Population Dynamics, Journal of Mathematical Biology, Vol.63, N.3, 519-556; [DOI 10.1007/s00285-010-0378-0].
- Caceres, M.O. and Caceres-Saez, I. 2013. Calculating effective growth rate from random Leslie model: Application to incidental mortality analysis, Ecological Modelling, 251, 312-322; [DOI: 10.1016/j.ecolmodel.2012.12.021]
- ↑ Hal Caswell (2001). Matrix Population Models: Construction, Analysis, and Interpretation. Sinauer.
Sources
- Krebs CJ (2001) Ecology: the experimental analysis of distribution and abundance (5th edition). San Francisco. Benjamin Cummings.
- Charlesworth, B. (1980) Evolution in age-structured population. Cambridge. Cambridge University Press
- Leslie, P.H. (1945) "The use of matrices in certain population mathematics". Biometrika, 33(3), 183–212.
- Leslie, P.H. (1948) "Some further notes on the use of matrices in population mathematics". Biometrika, 35(3–4), 213–245.
- Lotka, A.J. (1956) Elements of mathematical biology. New York. Dover Publications Inc.
- Kot, M. (2001) Elements of Mathematical Ecology, Cambridge. Cambridge University Press.