Giardia lamblia
| Giardia lamblia | |
|---|---|
 | |
| Giardia lamblia cell, SEM | |
| Scientific classification | |
| Domain: | Eukaryota |
| (unranked): | Excavata |
| Phylum: | Metamonada |
| Order: | Diplomonadida |
| Family: | Hexamitidae |
| Genus: | Giardia |
| Species: | G. lamblia |
| Binomial name | |
| Giardia lamblia (Lambl, 1859) Kofoid & Christiansen, 1915 | |
| Synonyms | |
| |
Giardia lamblia is a flagellated protozoan parasite that colonizes and reproduces in the small intestine, causing giardiasis. The parasite attaches to the epithelium by a ventral adhesive disc, and reproduces via binary fission.[1] Giardiasis does not spread via the bloodstream, nor does it spread to other parts of the gastrointestinal tract, but remains confined to the lumen of the small intestine.[2] Giardia trophozoites absorb their nutrients from the lumen of the small intestine, and are anaerobes. If the organism is split and stained, its characteristic pattern resembles the familiar "smiley face" symbol.[3] Chief pathways of human infection include ingestion of untreated sewage, a phenomenon particularly common in many developing countries;[4] contamination of natural waters also occurs in watersheds where intensive grazing occurs.
Hosts
Giardia infects humans, but is also one of the most common parasites infecting cats, dogs and birds. Mammalian hosts also include cattle, beavers, deer, and sheep.
Transmission
Giardia infection can occur through ingestion of dormant microbial cysts in contaminated water, food, or by the fecal-oral route (through poor hygiene practices). The cyst can survive for weeks to months in cold water,[5] so can be present in contaminated wells and water systems, especially stagnant water sources, such as naturally occurring ponds, storm water storage systems, and even clean-looking mountain streams. They may also occur in city reservoirs and persist after water treatment, as the cysts are resistant to conventional water treatment methods, such as chlorination and ozonolysis.[5] Zoonotic transmission is also possible, so Giardia infection is a concern for people camping in the wilderness or swimming in contaminated streams or lakes, especially the artificial lakes formed by beaver dams (hence the popular name for giardiasis, "beaver fever").
In addition to waterborne sources, fecal-oral transmission can also occur, for example in day-care centers, where children may have poor hygiene practices. Those who work with children are also at risk of being infected, as are family members of infected individuals. Not all Giardia infections are symptomatic, and many people can unknowingly serve as carriers of the parasite.
Life cycle
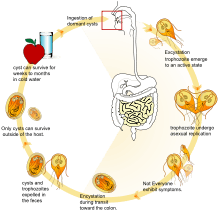
The life cycle begins with a noninfective cyst being excreted with the faeces of an infected individual. The cyst is hardy, providing protection from various degrees of heat and cold, desiccation, and infection from other organisms. A distinguishing characteristic of the cyst is four nuclei and a retracted cytoplasm. Once ingested by a host, the trophozoite emerges to an active state of feeding and motility. After the feeding stage, the trophozoite undergoes asexual replication through longitudinal binary fission. The resulting trophozoites and cysts then pass through the digestive system in the faeces. While the trophozoites may be found in the faeces, only the cysts are capable of surviving outside of the host.
Distinguishing features of the trophozoites are large karyosomes and lack of peripheral chromatin, giving the two nuclei a halo appearance. Cysts are distinguished by a retracted cytoplasm. This protozoan lacks mitochondria, although the discovery of the presence of mitochodrial remnants (organelles) in one recent study "indicate that Giardia is not primitively amitochondrial and that it has retained a functional organelle derived from the original mitochondrial endosymbiont".[6] This organelle is now termed a mitosome.
Intracellular metabolism and biochemistry
Giardia relies on glucose as its major energy source and breaks glucose down into ethanol, acetate and carbon dioxide.[7] However, it can also use arginine as an energy source.[8] Giardia possesses unique biochemical pathways that suggest it diverged from other eukaryotes at an early stage in evolution.[8]
B vitamins and bile salts, as well as glucose, are necessary for Giardia to survive, and a low-carbohydrate diet was shown in mice to reduce the number of Giardia organisms present.[9]
Microscopy

(A) is the cyst imaged by transmission (differential interference contrast), only.
(B) is the cyst wall selectively imaged through use of fluorescent-labelled (TRITC) antibody that is cyst-wall specific.
(C) is the cyst imaged through use of carboxy fluorescein diacetate, a viability stain.
(D) is a composite image of (B) and (C).
(E) is a composite image of (A), (B), and (C).
Under a normal compound light microscope, Giardia often looks like a "clown face," with two nuclei outlined by adhesive discs above dark median bodies that form the "mouth." Cysts are oval, have four nuclei, and have clearly visible axostyles. Most eukaryotes have mitochondria, though Giardia is one of the few to lack these, they do contain mitochondrial relics, called mitosomes.[6] Another such organism is Mixotricha paradoxa. However, similar to other eukaryotes, Giardia contains a complex endomembrane system as well as a tubulin based cytoskeleton.[10][11] Immunoelectron microscopy with antibody to alpha-tubulin shows that microtubules are present in Giardia in different structures such as axonemes, basal bodies, adhesive discs, funis and the median bodies.[11]
Research
Giardia alternates between two different forms — a hardy, dormant cyst that contaminates water or food, and an active, disease-causing form that emerges after the parasite is ingested. Dr. Frances Gillin of the University of California, San Diego and her colleagues cultivated the entire life cycle of this parasite in the laboratory, and identified biochemical cues in the host's digestive system which trigger Giardia's life cycle transformations.[12][13] They also uncovered several ways in which the parasite evades the defences of the infected organism. One of these is by altering the proteins on its surface, which confounds the ability of the infected animal's immune system to detect and combat the parasite (called antigenic variation). Gillin's work reveals why Giardia infections are extremely persistent and prone to recur. In addition, these insights into its biology and survival techniques may enable scientists to develop better strategies to understand, prevent, and treat Giardia infections.
In December 2008, Nature published an article showing the discovery of an RNA interference mechanism that allows Giardia to switch variant-specific surface proteins to avoid host immune response. The discovery was made by the team working at the Biochemistry and Molecular Biology Laboratory, School of Medicine, Catholic University of Cordoba, Argentina, led by Dr. Hugo Lujan.
Genomics
Giardia and the other diplomonads are unique in their possession of two nuclei that are similar in appearance, DNA content, transcription and time of replication. There are five chromosomes per the haploid genome. The genome has been sequenced and was published in 2007, although the sequence contains several gaps. The sequence is about 12 million base pairs and contains about 5000 protein-coding genes.[14] The GC content is 46%. Trophozoites have a ploidy of four and the ploidy of cysts is eight, which in turn raises the question of how Giardia maintains homogeneity between the chromosomes of the same and opposite nuclei. Modern sequencing technologies have been used to resequence different strains.[15]
Giardia had been assumed to be primitively asexual and with no means of transferring DNA between nuclei. These assumptions made it very difficult to explain the remarkably low level of allelic heterozygosity (< 0.01%) in the genome isolate, WB. However, all those assumptions of asexuality are now in doubt, with population genetics providing evidence for recombination[16] and the identification of meiotic genes, evidence for recombination among isolates and the evidence for exchange of genetic material between nuclei during the process of encystation.[17]
These findings on sexuality in Giardia, above, have important implications for understanding the origin of sexual reproduction in eukaryotes. Even though sexual reproduction is widespread among extant eukaryotes, it seemed unlikely, until recently, that sex is a primordial and fundamental feature of eukaryotes. A probable reason for the view that sex may not be fundamental to eukaryotes was that sexual reproduction previously appeared to be lacking in certain human pathogenic single-celled eukaryotes (e.g. Giardia) that diverged from early ancestors in the eukaryotic lineage.
In addition to the evidence cited above for recombination in Giardia, Malik et al.[18] reported that many meiosis specific genes occur in the Giardia genome, and further that homologs of these genes also occur in another unicellular eukaryote, Trichomonas vaginalis. Because these two species are descendants of lineages that are highly divergent among eukaryotes, Malik et al.[18] suggested that these meiotic genes were present in a common ancestor of all eukaryotes. Thus, on this view, the earliest ancestor of eukaryotes was likely capable of sexual reproduction. Furthermore, Dacks and Roger [19] proposed, based on phylogenetic analysis, that facultative sex was present in the common ancestor of all eukaryotes. Bernstein et al. also reviewed evidence in support of this view.[20]
Seven genotypes of Giardia have been recognized to date (A-G). Of these, B is the most widespread. Only types A and B have been shown to be infectious to humans.
History
The trophozoite form of Giardia was first observed in 1681 by Antonie van Leeuwenhoek in his own diarrhea stools. The genus was chosen to honour Professor Alfred Mathieu Giard of Paris.
The names for the human parasite Giardia duodenalis, Giardia lamblia and Giardia intestinalis are all in common current use despite the potential for confusion this has created.
Van Leeuwenhoek's observations were recreated, using a single-lens microscope of the kind he used, by British microbiologist Brian J. Ford, who showed how clearly one could view Giardia through a primitive microscope.[21]
In 1998, a highly publicised Giardia and Cryptosporidium outbreak was reported in Sydney, Australia, but it was found to be due to mismeasurement of the concentrations of microbes in the water supply. A 2004 outbreak in Bergen (Norway) hastened work on adding UV treatment to the water facilities.[22]
In October 2007, Giardia was found in the water supply for parts of Oslo, prompting authorities to advise the public to boil drinking water;[23] but subsequent test showed levels of contamination too low to pose a threat, so this advice has since been cancelled.[24]
In 2008, Giardia was identified as one of the causes of the dysentery afflicting Crusaders in Palestine in the 12th and 13th centuries.[25]
Other animals
Cats can be cured easily and lambs usually simply lose weight, but in calves, the parasites can be fatal and often are not responsive to antibiotics or electrolytes. Carriers among calves can also be asymptomatic. This parasite is deadly for chinchillas, so extra care must be taken by providing them with safe water. Dogs have a high infection rate, as 30% of the population under one year old are known to be infected in kennels. The infection is more prevalent in puppies than in adult dogs. Infected dogs can be isolated and treated, or the entire pack at a kennel can be treated together regardless. Kennels should also be then cleaned with bleach or other cleaning disinfectants. The grass areas used for exercise should be considered contaminated for at least one month after dogs show signs of infection, as cysts can survive in the environment for long periods of time. Prevention can be achieved by quarantine of infected dogs for at least 20 days and careful management and maintenance of a clean water supply.
See also
- Giardiasis
- List of parasites (human)
- 1998 Sydney water crisis
- Meiosis
- Origin and function of meiosis
References
- ↑ Oxford textbook of Medicine 1 (4th ed.). Oxford University Press. 2003. pp. 759–760. ISBN 0-19-262922-0.
- ↑ Harrison's Internal Medicine, Harrison's Online Chapter 199 Protozoal intestinal infections and trochomoniasis
- ↑ DeMay, Richard M. (1999). Practical principles of cytopathology. the University of Michigan: American Society for Clinical Pathology. p. 88. ISBN 9780891894377.
- ↑ Hogan, C. Michael (2010). "Water pollution". In McGinley, Mark; Cleveland, C. Encyclopedia of Earth. Washington DC: National Council for Science and the Environment.
- 1 2 Huang DB, White AC (2006). "An updated review on Cryptosporidium and Giardia". Gastroenterol. Clin. North Am. 35 (2): 291–314, viii. doi:10.1016/j.gtc.2006.03.006. PMID 16880067.
- 1 2 Tovar J; León-Avila G; Sánchez LB; et al. (2003). "Mitochondrial remnant organelles of Giardia function in iron-sulphur protein maturation". Nature 426 (6963): 172–6. doi:10.1038/nature01945. PMID 14614504.
- ↑ "Giardia — MicrobeWiki". Microbewiki.kenyon.edu. Retrieved 2010-07-29.
- 1 2 Brown DM, Upcroft JA, Edwards MR, Upcroft P (1998). "Anaerobic bacterial metabolism in the ancient eukaryote Giardia duodenalis". International Journal for Parasitology 28 (1): 149–64. doi:10.1016/S0020-7519(97)00172-0. PMID 9504342.
- ↑ Erlandsen; Meyer (1984). Giardia and Giardiasis. New York: Plenum Press. ISBN 0-306-41539-9.
- ↑ Soltys BJ, Falah M, Gupta RS (July 1996). "Identification of endoplasmic reticulum in the primitive eukaryote Giardia lamblia using cryoelectron microscopy and antibody to Bip". J. Cell. Sci. 109 (Pt 7): 1909–17. PMID 8832413.
- 1 2 Soltys BJ, Gupta RS (1994). "Immunoelectron microscopy of Giardia lamblia cytoskeleton using antibody to acetylated alpha-tubulin". J. Eukaryot. Microbiol. 41 (6): 625–32. doi:10.1111/j.1550-7408.1994.tb01524.x. PMID 7866386.
- ↑ Hetsko ML, McCaffery JM, Svärd SG, Meng TC, Que X, Gillin FD (1998). "Cellular and transcriptional changes during excystation of Giardia lamblia in vitro". Experimental Parasitology 88 (3): 172–83. doi:10.1006/expr.1998.4246. PMID 9562420.
- ↑ Svärd SG, Meng TC, Hetsko ML, McCaffery JM, Gillin FD (1998). "Differentiation-associated surface antigen variation in the ancient eukaryote Giardia lamblia". Molecular Microbiology 30 (5): 979–89. doi:10.1046/j.1365-2958.1998.01125.x. PMID 9988475.
- ↑ Morrison HG; McArthur AG; Gillin FD; et al. (2007). "Genomic minimalism in the early diverging intestinal parasite Giardia lamblia". Science 317 (5846): 1921–6. doi:10.1126/science.1143837. PMID 17901334.
- ↑ Franzén O; Jerlström-Hultqvist J; Castro E; et al. (2009). Petri, William, ed. "Draft Genome Sequencing of Giardia intestinalis Assemblage B Isolate GS: Is Human Giardiasis Caused by Two Different Species?". PLoS Pathogens 5 (8): e1000560. doi:10.1371/journal.ppat.1000560. PMC 2723961. PMID 19696920.
- ↑ Cooper MA, Adam RD, Worobey M, Sterling CR (November 2007). "Population genetics provides evidence for recombination in Giardia". Curr. Biol. 17 (22): 1984–8. doi:10.1016/j.cub.2007.10.020. PMID 17980591.
- ↑ Adam, RD; Svard, SG (2010). "Giardia: Nuclear and Chromosomal Structure and Replication". Anaerobic Parasitic Protozoa: Genomics and Molecular Biology. Caister Academic Press. ISBN 978-1-904455-61-5.
- 1 2 Malik SB, Pightling AW, Stefaniak LM, Schurko AM, Logsdon JM (2008). "An expanded inventory of conserved meiotic genes provides evidence for sex in Trichomonas vaginalis". PLoS ONE 3 (8): e2879. doi:10.1371/journal.pone.0002879. PMC 2488364. PMID 18663385.
- ↑ Dacks J, Roger AJ (June 1999). "The first sexual lineage and the relevance of facultative sex". J. Mol. Evol. 48 (6): 779–83. doi:10.1007/pl00013156. PMID 10229582.
- ↑ Bernstein H, Bernstein C, Michod RE (2012). "Ch. 1: DNA repair as the primary adaptive function of sex in bacteria and eukaryotes". In Sakura Kimura; Sora Shimizu. DNA Repair: New Research. Hauppauge NY: Nova Science. pp. 1–49. ISBN 978-1-62100-808-8.
- ↑ Ford BJ (2005). "The discovery of Giardia" (pdf). The Microscope 53 (4): 148–153.
- ↑ Nygård K, Schimmer B, Søbstad Ø, Walde A, Tveit I, Langeland N, Hausken T, Aavitsland P. (2006). "A large community outbreak of waterborne giardiasis-delayed detection in a non-endemic urban area". BMC Public Health 6 (1): 141. doi:10.1186/1471-2458-6-141. PMC 1524744. PMID 16725025.
- ↑ AVJonathan Tisdall . "Oslo water unsafe — Aftenposten — News in English". Aftenposten.no. Retrieved 2010-07-29.
- ↑ "Hovedside — Vann- og avløpsetaten — Oslo kommune". Vann-og-avlopsetaten.oslo.kommune.no. Retrieved 2010-07-29.
- ↑ Mitchell, Piers D.; Stern, Eliezer; Tepper, Yotam (2008). "Dysentery in the crusader kingdom of Jerusalem: an ELISA analysis of two medieval latrines in the City of Acre (Israel)". Journal of Archaeological Science 35 (7): 1849–1853. doi:10.1016/j.jas.2007.11.017.
External links
| Wikimedia Commons has media related to Giardia lamblia. |
- Giardia lamblia image library
- GiardiaDB: The Giardia lamblia genome sequencing project
- Washington State Department of Health fact sheet on Giardia.
- Centers for Disease Control and Prevention (CDC) Giardia Information
- United States Environmental Protection Agency fact sheet on Giardia in water
- Giardia article at MicrobeWiki
- Video of Giardia Life Cycle
- Giardia and the Sierra Nevada
- http://diarrhea.emedtv.com/giardia-lamblia/giardia-lambia.html
- Prucca CG; Slavin I; Quiroga R; et al. (2008). "Antigenic variation in Giardia lamblia is regulated by RNA interference". Nature 456 (7223): 750–4. doi:10.1038/nature07585. PMID 19079052. Lay summary – The New York Times (15 December 2008).
- "Giardia intestinalis". NCBI Taxonomy Browser. 5741.
| ||||||||||||||||||||||||||||