Janzen–Connell hypothesis
The Janzen–Connell hypothesis is a widely accepted explanation for the maintenance of tree species biodiversity in tropical rainforests. It was published independently in the early 1970s by Daniel Janzen[1] and Joseph Connell.[2] According to their hypothesis, host-specific herbivores, pathogens, or other natural enemies (often referred to as predators[1]) make the areas near a parent tree (the seed producing tree) inhospitable for the survival of seedlings. These natural enemies are referred to as 'distance-responsive predators' if they kill seeds or seedlings near the parent tree, or 'density-dependent predators' if they kill seeds or seedlings where they are most abundant (which is typically near the parent tree[1]). Such predators can prevent any one species from dominating the landscape, because if that species is too common, there will be few safe places for its seedlings to survive. However, because the predators are host-specific (also called specialists), they will not harm other tree species. As a result, if a species becomes very rare, then more predator-free areas will become available, giving that species' seedlings a competitive advantage. This negative feedback allows the tree species to coexist, and can be classified as a stabilizing mechanism.
The Janzen-Connell hypothesis has been called a special case of keystone predation, predator partitioning or the pest pressure hypothesis.[3][4] The pest pressure hypothesis states that plant diversity is maintained by specialist natural enemies.[5] The Janzen-Connell hypothesis expands on this, by claiming that the natural enemies are not only specialists, but also are distance-responsive or density-responsive.[1]
This mechanism has been proposed as promoting diversity of forests as it promotes survival of a number of different plant species within one localized region. While previously thought to explain the high diversity of tropical forests in particular, subsequent research has demonstrated the applicability of the Janzen–Connell hypothesis in temperate settings as well. The Black Cherry is one such example of a temperate forest species whose growth patterns can still be explained by the Janzen–Connell hypothesis.
History
Daniel Janzen’s hypothesis
Daniel Janzen published his hypothesis in 1970 in The American Naturalist under the article "Herbivores and the Number of Tree Species in Tropical Forests."[1] His hypothesis was based on the observation that in tropical forests (when compared to temperate forests), there were few new adult trees in the immediate vicinity of their parent tree. He explained the low density of tropical trees and lack of "bunching" of tree types around parent trees for two reasons: (1) the number of seeds decline with distance from the parent tree and (2) that the adult tree, its seeds, and seedlings are a source of food for host-specific parasites and diseases. Using his observations, Janzen created a model demonstrating the probability of a seed maturation or a seedling survival as a function of distance from the parent tree (as well as total seed count, dispersal mechanism, and predatorial activity).
Joseph Connell hypothesis
Joseph Connell published his hypothesis in 1971 in Dynamics of Populations.[2] Unlike Janzen, Connell proposed experiments that focused on the key prediction that exclusion of host-specific predators would cause a decrease in diversity as tree species with greater establishment or competitive ability formed low-diversity seedling and sapling communities where dominance was concentrated in a few species.
He formed his hypothesis through observations in Queensland, Australia. Along with Jack Greening Tracey and Larry Johnson Webb, he mapped trees in two rainforests and observed that smaller seedlings tended to occur in single-species clumps. Smaller seedlings also exhibited greater mortality, especially when their nearest neighbor was an individual of the same species. This pattern lessened with growth and age until seedlings exhibited similar pattern diversity to adults. To reinforce these observations, Connell ran an experiment showing that adult trees have a deleterious effect on smaller trees of the same species. In another experiment, Connell found that pre-germination predation was greater on seeds near adults of the same species than those near adults of others. Through these observations, Connell suggests that each tree species has host-specific enemies that attack it and any of its offspring which are close to the parent. This emphasizes the importance of the role of predation in preventing trees from forming single-species groves, which is probably the only way in which one species of tree could exclude others by interspecies competition.
Effects on forest dynamics
Disease dynamics and tree density
Plant pathogens follow infectious disease dynamics. The basic reproductive rate 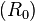 of a disease is dependent on three variables such that:
of a disease is dependent on three variables such that:
R0 = βL'S
Where β is the transmission rate or infectiousness of the disease, L is the average infection time of the host, and S is the density of the host population. By decreasing any one of the variables, the reproduction rate of the disease decreases. Since seed dispersal is such that the highest density of seeds is around the parent with density decreasing with distance from the parent, the reproduction rate of a disease infecting seeds and seedlings will be highest around the parent and decrease with distance. Thus, seedlings close to the parent are likely to die due to the disease prevalence. However, seedlings farther away are less likely to encounter the disease and therefore will more likely grow into adults.
Herbivory and tree density
Specialist herbivores who consume plant matter can also be thought of as having a "transmission rate" between individuals similarly to a disease. Tree predators (especially herbivorous insects) are limited by the ease of movement. When individuals are closer together at high density, movement between trees is easier and the predators quickly spread out. However, at low tree density, predators can not find the next individual with as much ease and thus often have low transmission rates leading to less specialist predation.
Concerns with the Janzen–Connell hypothesis
Many studies examining the Janzen–Connell hypothesis have shown supporting patterns with a number of tree species, but despite this there are also problematic aspects of the hypothesis.
- The first is that the Janzen–Connell hypothesis explains diversity at a community-wide scale, but most studies have only looked at a single species or a localized region. Most studies do not test the diversity prediction and did not determine the causes for patterns consistent with Janzen–Connell effects.[3] As Wright (2002) pointed out, "field measurements only demonstrate that niche differences, Janzen–Connell effects, and negative density dependence occur. Implications for species coexistence and plant diversity remain conjectural."[6]
- While these host-specific predators may play keystone species roles as they possibly aid in the prevention of superior tree competitors from monopolizing an area, no study has yet to examine species richness and abundance after removing natural enemies, creating a gap in the supporting research for the hypothesis.[3]
- Diversity may be maintained, at least in part, by episodic outbreaks of specialized pests, which may reduce the survivorship, growth, and reproductive success of adults of a species whenever they are particularly aggregated. Thus, specialist predation also impacts adult density, rather than just juveniles which has been the focus of research efforts.[3]
- Disease and predation may be just affecting overall density rather than in localized regions around adults alone.[3]
- Theoretical work has suggested that distance-responsive predators are less able to promote coexistence than equivalent predators that are not distance-responsive.[7]
- Many empirical studies have shown that few natural enemies are host-specific specialists,[8] as the original hypothesis claimed. However, theoretical work has suggested that predators can have broader diets as long as they are more damaging to their primary host than other species.[9]
Research testing the hypothesis
Supporting research
There have been over 50 studies designed to test predictions of this hypothesis in tropical and other settings and most show that many tree species exhibit patterns consistent with Janzen–Connell effects. Studies that have supported the Janzen–Connell hypothesis:
- A 2010 study of Panama forest reinforced that Janzen–Connell effects are also prevalent in tropical rainforests as seedlings growing close to their parents are likely to die due to microorganisms in the soil.[10]
- A 2008 study by Petermann et al. in grasslands showed that soil-borne pests created a feedback very similar to the Janzen–Connell effect, supporting the hypothesis as a driver of diversity in temperate ecosystems. This study suggests that the predator/density mechanism that promotes species diversity is not contained to tropical forests alone – even if tropical ecosystems do have the highest diversity. This supports the hypothesis as a mechanism for diversity, but not as an explanation latitudinal gradients in species diversity.[11]
- An examination of spatial data for 24 woody tropical rain forest plants showed either density-dependence or distance-dependence in plant offspring, supporting the hypothesis. However, other causal factors such as allelopathy were also suggested in some cases.[12]
Dissenting Research
Studies questioning the Janzen–Connell hypothesis:
- Hyatt et al. examined the hypothesis across a number of tree species (rather than focusing on the effects on just one) and found no support for the distance-dependence of the hypothesis and concluded that the Janzen–Connell hypothesis was not a mechanism for diversity. They did find however that in temperate settings, distance from parent reduced survivorship. But in tropical settings there was a slight positive correlation between the improved competitive ability of seeds and distance from parent.[13]
- A 1994 study by Burkey found that seed predation did not follow a pattern supporting that of the Janzen–Connell hypothesis along a meaningful scale. Seeds within 1 metre from the trunk were highly predated. However, seeds reached their peak density while still under the canopy of their parent. The authors concluded that seed predation did not follow the Janzen–Connell hypothesis.[14]
Current conclusions and further research
It is tricky to form conclusions regarding the accuracy of the Janzen–Connell hypothesis as it is difficult to falsify. This is because:
- Heavy predation may keep some species rare and widely spaced, and these species may also be the best competitors. If so, these species are most likely to form dense aggregations that would reduce diversity in local areas. But due to their rarity or a belief that rare species are not regulated by density-dependence, these species may be the ones least likely to be studied.[3]
- This means that the failure to find Janzen–Connell effects for what could be hundreds of tree species does not reject the hypothesis, as ecologists are missing the few key species where it does apply. However, dissenting findings do reduce the importance of the hypothesis for explaining overall coexistence of a number species.[3]
- For example, Hyatt et al. (2003) found that there were "individual cases of conformity to the hypothesis,"[13] which is all that is needed for the hypothesis to work if the specific cases represent tree species that are excellent competitors, highly shade tolerant, habitat generalists, or some combination of these traits that would allow these species to otherwise dominate the ecosystem.[3]
It is likely that a number of mechanisms underscore the coexistence of similar species and thus cause biodiversity in ecosystems. It is possible the Janzen–Connell hypothesis is applicable only for some species depending on species characteristics. The hypothesis may also be affected by the kind of predator or pathogen as preliminary research has shown that the hypothesis is true only when host-specific predators have limited mobility with a range less than the seed dispersal range.[15]
References
- 1 2 3 4 5 Janzen, Daniel H. "Herbivores and the Number of Tree Species in Tropical Forests." The American Naturalist. 104.940 (1970).
- 1 2 Connell, J.H. "On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees." In: Dynamics of Population. Ed. P.J. Den Boer and G.R. Gradwell. Wageningen: Pudoc, 1970.
- 1 2 3 4 5 6 7 8 Carson, Walter A. and Stefan A. Schnitzer. Tropical Forest Community Ecology. Blackwell Publishing: Hoboken, NJ, 2008.
- ↑ Gilbert, Gregory (2005). Burslem, David; Pinard, Michelle; Hartley, Sue, eds. Biotic interactions in the tropics : their role in the maintenance of species diversity. Cambridge, UK: Cambridge University Press. pp. 141–164. ISBN 9780521609852.
- ↑ Gillette, J.B. (1962). "Pest pressure, an underestimated factor in evolution". Taxonomy and Geography; a symposium 4 (37): 37–46.
- ↑ Wright, J.S. "Plant diversity in tropical forests: a review of mechanisms of species coexistence." Oecologia. 130.1 (2002): 1–14.
- ↑ Stump, Simon Maccracken; Chesson, Peter (December 2015). "Distance-responsive predation is not necessary for the Janzen–Connell hypothesis". Theoretical Population Biology 106: 60–70. doi:10.1016/j.tpb.2015.10.006.
- ↑ Novotny, V.; Basset, Y. (7 June 2005). "Host specificity of insect herbivores in tropical forests". Proceedings of the Royal Society B: Biological Sciences 272 (1568): 1083–1090. doi:10.1098/rspb.2004.3023.
- ↑ Sedio, Brian E.; Ostling, Annette M.; Ris Lambers, Janneke Hille (August 2013). "How specialised must natural enemies be to facilitate coexistence among plants?". Ecology Letters 16 (8): 995–1003. doi:10.1111/ele.12130.
- ↑ Mangan, Scott A., et al. "Negative plant–soil feedback predicts tree-species relative abundance in a tropical forest." Nature 466 (8 July 2010) 752–755.
- ↑ Pertermann, Jana S. et al. "Janzen–Connell Effects are Widespread and Strong Enough to Maintain Diversity in Grasslands." Ecology 89.9 (2008): 2399–2406.
- ↑ Clark, Deborah A. and David B. Clark. "Dynamics of a Tropical Rain Forest Tree: Evaluation of the Janzen–Connell Model." The American Naturalist. 124.6 (1984):769–788.
- 1 2 Hyatt, Lauren A. et al. "The Distance Dependence Prediction of the Janzen–Connell Hypothesis: a Meta-Analysis." Oikos. 103.3 (2003): 590–602.
- ↑ Burkey, T.V. "Tree Species Diversity: A Test of the Janzen–Connell Model." Oecologia. 97. (1994): 533–540.
- ↑ Nathan, Ran and Renato Casagrandi. "A simple mechanistic model of seed dispersal, predation and plant establishment: Janzen–Connell and beyond." Journal of Ecology. 92. (2004): 733–746.