Human serum albumin
| human serum albumin | |
|---|---|
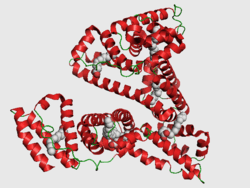 PDB rendering based on 1e7h. | |
| Identifiers | |
| Symbol | ALB |
| Entrez | 213 |
| HUGO | 399 |
| OMIM | 103600 |
| PDB | 1E7H |
| RefSeq | NM_000477 |
| UniProt | P02768 |
| Other data | |
| Locus | Chr. 4 q13.3 |
Human serum albumin is the version of serum albumin found in human blood. It is the most abundant protein in human blood plasma; it constitutes about half of serum protein. It is produced in the liver. It is soluble and monomeric.
Albumin transports hormones, fatty acids, and other compounds, buffers pH, and maintains oncotic pressure, among other functions.
Albumin is synthesized in the liver as preproalbumin, which has an N-terminal peptide that is removed before the nascent protein is released from the rough endoplasmic reticulum. The product, proalbumin, is in turn cleaved in the Golgi vesicles to produce the secreted albumin.
The reference range for albumin concentrations in serum is approximately 35 - 50 g/L (3.5 - 5.0 g/dL).[1] It has a serum half-life of approximately 20 days. It has a molecular mass of 66.5 kDa.
The gene for albumin is located on chromosome 4 and mutations in this gene can result in anomalous proteins. The human albumin gene is 16,961 nucleotides long from the putative 'cap' site to the first poly(A) addition site. It is split into 15 exons that are symmetrically placed within the 3 domains thought to have arisen by triplication of a single primordial domain.
Function
- Maintains oncotic pressure
- Transports thyroid hormones
- Transports other hormones, in particular, ones that are fat-soluble
- Transports fatty acids ("free" fatty acids) to the liver and to myocytes for utilization of energy
- Transports unconjugated bilirubin
- Transports many drugs; serum albumin levels can affect the half-life of drugs
- Competitively binds calcium ions (Ca2+)
- Buffers pH
- Serum albumin, as a negative acute-phase protein, is down-regulated in inflammatory states. As such, it is not a valid marker of nutritional status; rather, it is a marker of an inflammatory state
- Prevents photodegradation of folic acid
Measurement
Serum albumin is commonly measured by recording the change in absorbance upon binding to a dye such as bromocresol green or bromocresol purple.[2]
Reference ranges
Serum albumin concentration is typically 35 - 50 g/L (3.5 - 5.0 g/dL)[1]
Pathology
Hypoalbuminemia
Hypoalbuminemia is a low blood albumin levels.[3] This can be caused by:
- Liver disease; cirrhosis of the liver is most common
- Excess excretion by the kidneys (as in nephrotic syndrome)
- Excess loss in bowel (protein-losing enteropathy, e.g., Ménétrier's disease)
- Burns (plasma loss in the absence of skin barrier)
- Redistribution (hemodilution [as in pregnancy], increased vascular permeability or decreased lymphatic clearance)
- Acute disease states (referred to as a negative acute-phase protein)
- Malnutrition and wasting[4]
- Mutation causing analbuminemia (very rare)
Hyperalbuminemia
Hyperalbuminemia is an increased concentration of albumin in the blood.[5] Typically, this condition is due to abrupt dehydration.[5]
Chronic dehydration needs to be treated with zinc as well as with water. Zinc reduces cell swelling caused by decreased intake of water (hypotonicity) and also increases retention of salt. In the dehydrated state, the body has too high an osmolarity and, it appears, discards zinc to prevent this. Zinc also regulates transport of the cellular osmolyte taurine, and albumin is known to increase cellular taurine absorption. Zinc has been shown to increase retinol (vitamin A) production from beta-carotene, and in lab experiments retinol reduced human albumin production.[6] It is possible that a retinol (vitamin A) deficiency alone could cause albumin levels to become raised. Patients recovering from chronic dehydration may develop dry eyes as the body uses up its vitamin A store. Retinol causes cells to swell with water (this is most likely one reason that too much vitamin A is toxic).[7] Hyperalbuminemia is also associated with high protein diets.[8]
Therapeutic uses
Human albumin solution or HSA is available for medical use, usually at concentrations of 5-25%.
Human albumin is often used to replace lost fluid and help restore blood volume in trauma, burns and surgery patients. A Cochrane systematic review[9] of 37 trials found no evidence that albumin, compared with cheaper alternatives such as saline, reduces the risk of dying.
Human serum albumin has been used as a component of a frailty index.[4]
It has not been shown to give better results than other fluids when used simply to replace volume, but is frequently used in conditions where loss of albumin is a major problem, such as liver disease with ascites.
Glycation
It has been known for a long time that human blood proteins like hemoglobin[10] and serum albumin[11][12] may undergo a slow non-enzymatic glycation, mainly by formation of a Schiff base between ε-amino groups of lysine (and sometimes arginine) residues and glucose molecules in blood (Maillard reaction). This reaction can be inhibited in the presence of antioxidant agents.[13] Although this reaction may happen normally,[11] elevated glycoalbumin is observed in diabetes mellitus.[12]
Glycation has the potential to alter the biological structure and function of the serum albumin protein.[14][15][16][17]
Moreover, the glycation can result in the formation of Advanced Glycation End-Products (AGE), which result in abnormal biological effects. Accumulation of AGEs leads to tissue damage via alteration of the structures and functions of tissue proteins, stimulation of cellular responses, through receptors specific for AGE-proteins, and generation of reactive oxygen intermediates. AGEs also react with DNA, thus causing mutations and DNA transposition. Thermal processing of proteins and carbohydrates brings major changes in allergenicity. AGEs are antigenic and represent many of the important neoantigens found in cooked or stored foods.[18] They also interfere with the normal product of nitric oxide in cells.[19]
Although there are several lysine and arginine residues in the serum albumin structure, very few of them can take part in the glycation reaction.[12][20] It is not clear exactly why only these residues are glycated in serum albumin, but it is suggested that non-covalent binding of glucose to serum albumin prior to the covalent bond formation might be the reason.[21]
Loss via kidneys
In the healthy kidney, albumin's size and negative electric charge exclude it from excretion in the glomerulus. This is not always the case, as in some diseases including diabetic nephropathy, which can sometimes be a complication of uncontrolled or of longer term diabetes in which proteins can cross the glomerulus. The lost albumin can be detected by a simple urine test.[22] Depending on the amount of albumin lost, a patient may have normal renal function, microalbuminuria, or albuminuria.
Amino acid sequence
The approximate sequence of human serum albumin is:[23]
MKWVTFISLL FLFSSAYSRG VFRRDAHKSE VAHRFKDLGE ENFKALVLIA FAQYLQQCPF EDHVKLVNEV TEFAKTCVAD ESAENCDKSL HTLFGDKLCT VATLRETYGE MADCCAKQEP ERNECFLQHK DDNPNLPRLV RPEVDVMCTA FHDNEETFLK KYLYEIARRH PYFYAPELLF FAKRYKAAFT ECCQAADKAA CLLPKLDELR DEGKASSAKQ RLKCASLQKF GERAFKAWAV ARLSQRFPKA EFAEVSKLVT DLTKVHTECC HGDLLECADD RADLAKYICE NQDSISSKLK ECCEKPLLEK SHCIAEVEND EMPADLPSLA ADFVESKDVC KNYAEAKDVF LGMFLYEYAR RHPDYSVVLL LRLAKTYETT LEKCCAAADP HECYAKVFDE FKPLVEEPQN LIKQNCELFE QLGEYKFQNA LLVRYTKKVP QVSTPTLVEV SRNLGKVGSK CCKHPEAKRM PCAEDYLSVV LNQLCVLHEK TPVSDRVTKC CTESLVNRRP CFSALEVDET YVPKEFNAET FTFHADICTL SEKERQIKKQ TALVELVKHK PKATKEQLKA VMDDFAAFVE KCCKADDKET CFAEEGKKLV AASQAALGL
Of the 609 amino acids in this sequence, encoded by the ALB gene and translated to form the precursor protein, only 585 amino acids are observed in the final product present in the blood; the first 24 amino acids (here italicized), including the signal peptide (1–18) and propeptide (19–22, or 19–24) portions, are cleaved after translation.
Interactions
Human serum albumin has been shown to interact with FCGRT.[24]
See also
References
- 1 2 "Harmonisation of Reference Intervals" (PDF). pathologyharmony.co.uk. Pathology Harmony. Retrieved 23 June 2013.
- ↑ "Albumin: analyte monograph" (PDF). Association for Clinical Biochemistry and Laboratory Medicine. Retrieved 23 June 2013.
- ↑ Anderson, Douglas M. (2000). Dorland's illustrated medical dictionary (29. ed.). Philadelphia [u.a.]: Saunders. p. 860. ISBN 0721682618.
- 1 2 Green P, Woglom AE, Genereux P, Daneault B, Paradis JM, Schnell S, Hawkey M, Maurer MS, Kirtane AJ, Kodali S, Moses JW, Leon MB, Smith CR, Williams M (2012). "The impact of frailty status on survival after transcatheter aortic valve replacement in older adults with severe aortic stenosis: a single-center experience". JACC Cardiovascular Interventions 5 (9): 974–981. doi:10.1016/j.jcin.2012.06.011. PMC 3717525. PMID 22995885.
- 1 2 Walker, edited by H. Kenneth; Hall, W. Dallas; Schlossberg, J. Willis Hurst ; illustrations by Leon; Boyter, Charles H. (1990). Clinical methods : the history, physical, and laboratory examinations (3rd ed.). Boston: Butterworths. p. Chapter 101. ISBN 040990077X.
- ↑ Masaki T, Matsuura T, Ohkawa K, Miyamura T, Okazaki I, Watanabe T, Suzuki T (July 2006). "All-trans retinoic acid down-regulates human albumin gene expression through the induction of C/EBPβ-LIP". Biochem. J. 397 (2): 345–53. doi:10.1042/BJ20051863. PMC 1513275. PMID 16608438.
- ↑ Pasantes-Morales H, Wright CE, Gaull GE (December 1984). "Protective effect of taurine, zinc and tocopherol on retinol-induced damage in human lymphoblastoid cells". J. Nutr. 114 (12): 2256–61. PMID 6502269.
- ↑ Mutlu EA, Keshavarzian A, Mutlu GM (June 2006). "Hyperalbuminemia and elevated transaminases associated with high-protein diet". Scand. J. Gastroenterol. 41 (6): 759–60. doi:10.1080/00365520500442625. PMID 16716979.
- ↑ The Albumin Reviewers (Alderson P, Bunn F, Li Wan Po A, Li L, Pearson M, Roberts I, Schierhout G). Human albumin solution for resuscitation and volume expansion in critically ill patients" Cochrane Database of Systematic Reviews 2004, Issue 4. Art. No.: CD001208. doi:10.1002/14651858.CD001208.pub2.
- ↑ Rahbar S (October 1968). "An abnormal hemoglobin in red cells of diabetics". Clin. Chim. Acta 22 (2): 296–8. doi:10.1016/0009-8981(68)90372-0. PMID 5687098.
- 1 2 Day JF, Thorpe SR, Baynes JW (February 1979). "Nonenzymatically glucosylated albumin. In vitro preparation and isolation from normal human serum". J. Biol. Chem. 254 (3): 595–7. PMID 762083.
- 1 2 3 Iberg N, Flückiger R (October 1986). "Nonenzymatic glycosylation of albumin in vivo. Identification of multiple glycosylated sites". J. Biol. Chem. 261 (29): 13542–5. PMID 3759977.
- ↑ Jakus V, Hrnciarová M, Cársky J, Krahulec B, Rietbrock N (1999). "Inhibition of nonenzymatic protein glycation and lipid peroxidation by drugs with antioxidant activity". Life Sci. 65 (18–19): 1991–3. doi:10.1016/S0024-3205(99)00462-2. PMID 10576452.
- ↑ Mohamadi-Nejad A, Moosavi-Movahedi AA, Hakimelahi GH, Sheibani N (September 2002). "Thermodynamic analysis of human serum albumin interactions with glucose: insights into the diabetic range of glucose concentration". Int. J. Biochem. Cell Biol. 34 (9): 1115–24. doi:10.1016/S1357-2725(02)00031-6. PMID 12009306.
- ↑ Shaklai N, Garlick RL, Bunn HF (March 1984). "Nonenzymatic glycosylation of human serum albumin alters its conformation and function". J. Biol. Chem. 259 (6): 3812–7. PMID 6706980.
- ↑ Mendez DL, Jensen RA, McElroy LA, Pena JM, Esquerra RM (December 2005). "The effect of non-enzymatic glycation on the unfolding of human serum albumin". Arch. Biochem. Biophys. 444 (2): 92–9. doi:10.1016/j.abb.2005.10.019. PMID 16309624.
- ↑ Mohamadi-Nejada A, Moosavi-Movahedi AA,Safariana S, Naderi-Maneshc MH, Ranjbarc B, Farzamid B, Mostafavie H, Larijanif MB, Hakimelahi GH, A (July 2002). "The thermal analysis of nonezymatic glycosylation of human serum albumin: differential scanning calorimetry and circular dichroism studies". Thermochimica Acta 389 (1–2): 141–151. doi:10.1016/S0040-6031(02)00006-0.
- ↑ Kańska U, Boratyński J (2002). "Thermal glycation of proteins by D-glucose and D-fructose". Arch. Immunol. Ther. Exp. (Warsz.) 50 (1): 61–6. PMID 11916310.
- ↑ Rojas A, Romay S, González D, Herrera B, Delgado R, Otero K (February 2000). "Regulation of endothelial nitric oxide synthase expression by albumin-derived advanced glycosylation end products". Circ. Res. 86 (3): E50–4. doi:10.1161/01.RES.86.3.e50. PMID 10679490.
- ↑ Garlick RL, Mazer JS (May 1983). "The principal site of nonenzymatic glycosylation of human serum albumin in vivo". J. Biol. Chem. 258 (10): 6142–6. PMID 6853480.
- ↑ Marashi SA, Safarian S, Moosavi-Movahedi AA (2005). "Why major nonenzymatic glycation sites of human serum albumin are preferred to other residues?". Med. Hypotheses 64 (4): 881. doi:10.1016/j.mehy.2004.11.007. PMID 15694713.
- ↑ Microalbumin Urine Test
- ↑ P02768
- ↑ Chaudhury C, Mehnaz S, Robinson JM, Hayton WL, Pearl DK, Roopenian DC, Anderson CL (February 2003). "The Major Histocompatibility Complex–related Fc Receptor for IgG (FcRn) Binds Albumin and Prolongs Its Lifespan". J. Exp. Med. 197 (3): 315–22. doi:10.1084/jem.20021829. PMC 2193842. PMID 12566415.
Further reading
- Komatsu T, Nakagawa A, Curry S, Tsuchida E, Murata K, Nakamura N, Ohno H (2009). "The role of an amino acid triad at the entrance of the heme pocket in human serum albumin for O(2) and CO binding to iron protoporphyrin IX". Org. Biomol. Chem. 7 (18): 3836–41. doi:10.1039/b909794e. PMID 19707690.
- Milojevic J, Raditsis A, Melacini G (2009). "Human Serum Albumin Inhibits Aβ Fibrillization through a "Monomer-Competitor" Mechanism". Biophys. J. 97 (9): 2585–94. doi:10.1016/j.bpj.2009.08.028. PMC 2770600. PMID 19883602.
- Silva AM, Hider RC (2009). "Influence of non-enzymatic post-translation modifications on the ability of human serum albumin to bind iron. Implications for non-transferrin-bound iron speciation". Biochim. Biophys. Acta 1794 (10): 1449–58. doi:10.1016/j.bbapap.2009.06.003. PMID 19505594.
- Otosu T, Nishimoto E, Yamashita S (2010). "Multiple conformational state of human serum albumin around single tryptophan residue at various pH revealed by time-resolved fluorescence spectroscopy". J. Biochem. 147 (2): 191–200. doi:10.1093/jb/mvp175. PMID 19884191.
- Blindauer CA, Harvey I, Bunyan KE, Stewart AJ, Sleep D, Harrison DJ, Berezenko S, Sadler PJ (2009). "Structure, Properties, and Engineering of the Major Zinc Binding Site on Human Albumin". J. Biol. Chem. 284 (34): 23116–24. doi:10.1074/jbc.M109.003459. PMC 2755717. PMID 19520864.
- Juárez J, López SG, Cambón A, Taboada P, Mosquera V (2009). "Influence of electrostatic interactions on the fibrillation process of human serum albumin". J Phys Chem B 113 (30): 10521–9. doi:10.1021/jp902224d. PMID 19572666.
- Fu BL, Guo ZJ, Tian JW, Liu ZQ, Cao W (2009). "[Advanced glycation end products induce expression of PAI-1 in cultured human proximal tubular epithelial cells through NADPH oxidase dependent pathway]". Xi Bao Yu Fen Zi Mian Yi Xue Za Zhi 25 (8): 674–7. PMID 19664386.
- Ascenzi P, di Masi A, Coletta M, Ciaccio C, Fanali G, Nicoletti FP, Smulevich G, Fasano M (2009). "Ibuprofen Impairs Allosterically Peroxynitrite Isomerization by Ferric Human Serum Heme-Albumin". J. Biol. Chem. 284 (45): 31006–17. doi:10.1074/jbc.M109.010736. PMC 2781501. PMID 19734142.
- Sowa ME, Bennett EJ, Gygi SP, Harper JW (2009). "Defining the Human Deubiquitinating Enzyme Interaction Landscape". Cell 138 (2): 389–403. doi:10.1016/j.cell.2009.04.042. PMC 2716422. PMID 19615732.
- Curry S (2002). "Beyond expansion: structural studies on the transport roles of human serum albumin". Vox Sang. 83 Suppl 1: 315–9. doi:10.1111/j.1423-0410.2002.tb05326.x. PMID 12617161.
- Guo S, Shi X, Yang F, Chen L, Meehan EJ, Bian C, Huang M (2009). "Structural basis of transport of lysophospholipids by human serum albumin". Biochem. J. 423 (1): 23–30. doi:10.1042/BJ20090913. PMID 19601929.
- de Jong PE, Gansevoort RT (2009). "Focus on microalbuminuria to improve cardiac and renal protection". Nephron Clin Pract 111 (3): c204–10; discussion c211. doi:10.1159/000201568. PMID 19212124.
- Page TA, Kraut ND, Page PM, Baker GA, Bright FV (2009). "Dynamics of loop 1 of domain I in human serum albumin when dissolved in ionic liquids". J Phys Chem B 113 (38): 12825–30. doi:10.1021/jp904475v. PMID 19711930.
- Roche M, Rondeau P, Singh NR, Tarnus E, Bourdon E (2008). "The antioxidant properties of serum albumin". FEBS Lett. 582 (13): 1783–7. doi:10.1016/j.febslet.2008.04.057. PMID 18474236.
- Wyatt AR, Wilson MR (2010). "Identification of Human Plasma Proteins as Major Clients for the Extracellular Chaperone Clusterin". J. Biol. Chem. 285 (6): 3532–9. doi:10.1074/jbc.M109.079566. PMC 2823492. PMID 19996109.
- Cui FL, Yan YH, Zhang QZ, Qu GR, Du J, Yao XJ (2010). "A study on the interaction between 5-Methyluridine and human serum albumin using fluorescence quenching method and molecular modeling". J Mol Model 16 (2): 255–62. doi:10.1007/s00894-009-0548-4. PMID 19588173.
- Caridi G, Dagnino M, Simundic AM, Miler M, Stancic V, Campagnoli M, Galliano M, Minchiotti L (2010). "Albumin Benkovac (c.1175 A > G; p.Glu392Gly): a novel genetic variant of human serum albumin". Translational Research 155 (3): 118–9. doi:10.1016/j.trsl.2009.10.001. PMID 20171595.
- Deeb O, Rosales-Hernández MC, Gómez-Castro C, Garduño-Juárez R, Correa-Basurto J (2010). "Exploration of human serum albumin binding sites by docking and molecular dynamics flexible ligand-protein interactions". Biopolymers 93 (2): 161–70. doi:10.1002/bip.21314. PMID 19785033.
- Karahan SC, Koramaz I, Altun G, Uçar U, Topbaş M, Menteşe A, Kopuz M (2010). "Ischemia-modified albumin reduction after coronary bypass surgery is associated with the cardioprotective efficacy of cold-blood cardioplegia enriched with N-acetylcysteine: a preliminary study". Eur Surg Res 44 (1): 30–6. doi:10.1159/000262324. PMID 19955769.
- Jin C, Lu L, Zhang RY, Zhang Q, Ding FH, Chen QJ, Shen WF (2009). "Association of serum glycated albumin, C-reactive protein and ICAM-1 levels with diffuse coronary artery disease in patients with type 2 diabetes mellitus". Clin. Chim. Acta 408 (1–2): 45–9. doi:10.1016/j.cca.2009.07.003. PMID 19615354.
External links
- Human Albumin structure in the Protein data bank
- Human Albumin information in the Swis-Prot/TrEMBL database
- Human Serum Albumin on the Human Protein Reference Database
- Albumin binding prediction
- Albumin at Lab Tests Online
- Albumin: analyte monograph from the Association for Clinical Biochemistry and Laboratory Medicine
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||









































{kind=link}
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| ||||||||||||||