Coalescent theory
Coalescent theory is a retrospective stochastic model of population genetics that relates genetic diversity in a sample to demographic history of the population from which it was taken. That is, it is a model of the effect of genetic drift, viewed backwards in time, on the genealogy of antecedents. It comprises a probabilistic assessment of variation in time to common ancestry of alleles in a relatively small sample of individuals, from a much larger population. This includes consideration of all pathways of inheritance through which sampled copies of a homologous DNA element are traced back to a single ancestral copy, known as the most recent common ancestor (MRCA; sometimes also termed the coancestor to emphasize the coalescent relationship). The inheritance relationships among alleles are typically represented as a gene genealogy, or gene tree, similar in form to a phylogenetic tree. The probabilistic expectation of this gene genealogy is also known as the coalescent. Understanding the statistical properties of the coalescent under different assumptions forms the basis of coalescent theory. Because of recombination, different gene loci follow different pathways of ancestry, resulting in different gene genealogies. The coalescent is also relevant to phylogenetics, as incomplete lineage sorting between speciation events results in conflict among gene-loci in phylogenetic relationships inferred among species.
The mathematical theory of the coalescent was originally developed in the early 1980s by John Kingman. In the simplest case, coalescent theory assumes no recombination, no natural selection, and no gene flow or population structure. The gene genealogy is independent of the mutational process, such that changes in the DNA sequence do not affect inheritance and can be considered separately (even if all gene copies are identical in sequence they are not equally related in the gene tree). Under this model, the expected time between successive coalescence events, by which two gene copies arise from a single ancestral copy, increases almost exponentially back in time (with wide variance). Advances in coalescent theory include recombination, selection, and virtually any arbitrarily complex evolutionary or demographic model in population genetic analysis.
Theory
Time to coalescence
Consider a single gene locus sampled from two haploid individuals in a population. The ancestry of this sample is traced backwards in time to the point where these two lineages coalesce in their MRCA. Coalescent theory seeks to estimate the expectation of this time period and its variance.
The probability that two lineages coalesce in the immediately preceding generation is the probability that they share a parental DNA sequence. In a population with a constant effective population size with 2Ne copies of each locus, there are 2Ne "potential parents" in the previous generation. Under a random mating model, the probability that two alleles originate from the same parental copy is thus 1/(2Ne) and, correspondingly, the probability that they do not coalesce is 1 − 1/(2Ne).
At each successive preceding generation, the probability of coalescence is geometrically distributed — that is, it is the probability of noncoalescence at the t − 1 preceding generations multiplied by the probability of coalescence at the generation of interest:
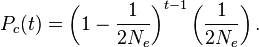
For sufficiently large values of Ne, this distribution is well approximated by the continuously defined exponential distribution

This is mathematically convenient as the standard exponential distribution has both the expected value and the standard deviation equal to 2Ne; therefore, although the expected time to coalescence is 2Ne, actual coalescence times have a wide range of variation. Note that coalescent time is the number of preceding generations where the coalescence took place and not calendar time though an estimation of the latter can be made multiplying 2Ne with the average time between generations. The above calculations apply equally to a diploid population of effective size Ne (in other words, for a non-recombining segment of DNA, each chromosome can be treated as equivalent to an independent haploid individual; in the absence of inbreeding, sister chromosomes in a single individual are no more closely related than two chromosomes randomly sampled from the population). Some effectively haploid DNA elements, such as mitochondrial DNA, however, are only carried by one sex and therefore have one quarter the effective size of the equivalent diploid population (Ne/2)
Neutral variation
Coalescent theory can also be used to model the amount of variation in DNA sequences expected from genetic drift and mutation. This value is termed the mean heterozygosity, represented as 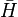 . Mean heterozygosity is calculated as the probability of a mutation occurring at a given generation divided by the probability of any "event" at that generation (either a mutation or a coalescence). The probability that the event is a mutation is the probability of a mutation in either of the two lineages:
. Mean heterozygosity is calculated as the probability of a mutation occurring at a given generation divided by the probability of any "event" at that generation (either a mutation or a coalescence). The probability that the event is a mutation is the probability of a mutation in either of the two lineages: 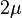 . Thus the mean heterozygosity is equal to
. Thus the mean heterozygosity is equal to
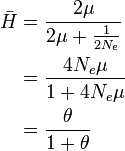
For 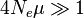 , the vast majority of allele pairs have at least one difference in nucleotide sequence.
, the vast majority of allele pairs have at least one difference in nucleotide sequence.
Graphical representation
Coalescents can be visualised using dendrograms which show the relationship of branches of the population to each other. The point where two branches meet indicates a coalescent event.
Applications
Disease gene mapping
The utility of coalescent theory in the mapping of disease is slowly gaining more appreciation; although the application of the theory is still in its infancy, there are a number of researchers who are actively developing algorithms for the analysis of human genetic data that utilise coalescent theory.
The genomic distribution of heterozygosity
The human single-nucleotide polymorphism (SNP) map has revealed large regional variations in heterozygosity, more so than can be explained on the basis of (Poisson-distributed) random chance. In part, these variations could be explained on the basis of assessment methods, the availability of genomic sequences, and possibly the standard coalescent population genetic model. Population genetic influences could have a major influence on this variation: some loci presumably would have comparatively recent common ancestors, others might have much older genealogies, and so the regional accumulation of SNPs over time could be quite different. The local density of SNPs along chromosomes appears to cluster in accordance with a variance to mean power law and to obey the Tweedie compound Poisson distribution. In this model the regional variations in the SNP map would be explained by the accumulation of multiple small genomic segments through recombination, where the mean number of SNPs per segment would be gamma distributed in proportion to a gamma distributed time to the most recent common ancestor for each segment.
History
Coalescent theory is a natural extension of the more classical population genetics concept of neutral evolution and is an approximation to the Fisher–Wright (or Wright–Fisher) model for large populations. It was discovered independently by several researchers in the 1980s.
Software
A large body of software exists for both simulating data sets under the coalescent process as well as inferring parameters such as population size and migration rates from genetic data.
- BEAST – Bayesian MCMC inference package with a wide range of coalescent models including the use of temporally sampled sequences.
- COAL – Program for computing gene tree probabilities and simulating gene trees in species trees under the coalescent model .
- CoaSim – software for simulating genetic data under the coalescent model.
- DIYABC - A user-friendly approach to ABC for inference on population history using molecular markers
- DendroPy - A Python library for phylogenetic computing, with classes and methods for simulating pure (unconstrained) coalescent trees as well as constrained coalescent trees under the multispecies coalescent model (i.e., "gene trees in species trees").
- GeneRecon – software for the fine-scale mapping of linkage disequilibrium mapping of disease genes using coalescent theory based on an Bayesian MCMC framework.
- genetree software for estimation of population genetics parameters using coalescent theory and simulation (the R package popgen). See also Oxford Mathematical Genetics and Bioinformatics Group
- GENOME – rapid coalescent-based whole-genome simulation
- IBDSim – A computer package for the simulation of genotypic data under general isolation by distance models .
- IMa – IMa implements the same Isolation with Migration model, but does so using a new method that provides estimates of the joint posterior probability density of the model parameters. IMa also allows log likelihood ratio tests of nested demographic models. IMa is based on a method described in Hey and Nielsen (2007 PNAS 104:2785–2790). IMa is faster and better than IM (i.e. by virtue of providing access to the joint posterior density function), and it can be used for most (but not all) of the situations and options that IM can be used for.
- Lamarc – software for estimation of rates of population growth, migration, and recombination.
- Migraine – A program which implements coalescent algorithms for a maximum likelihood analysis (using Importance Sampling algorithms) of genetic data with a focus on spatially structured populations .
- Migrate – Maximum likelihood and Bayesian inference of migration rates under the n-coalescent. The inference is implemented using MCMC
- MaCS – Markovian Coalescent Simulator - Simulates genealogies spatially across chromosomes as a Markovian process. Similar to the SMC algorithm of McVean and Cardin, and supports all demographic scenarios found in Hudson's ms.
- ms & msHOT – Richard Hudson's original program for generating samples under neutral models and an extension which allows recombination hotspots.
- msms – An extended version of ms that includes selective sweeps.
- Recodon and NetRecodon – software to simulate coding sequences with inter/intracodon recombination, migration, growth rate and longitudinal sampling .
- SARG – Structure Ancestral Recombination Graph by Magnus Nordborg
- simcoal2 – software to simulate genetic data under the coalescent model with complex demography and recombination
- TreesimJ Forward simulation software allowing sampling of genealogies and data sets under diverse selective and demographic models.
References and notes
Articles
- ^ Arenas, M. and Posada, D. (2007) Recodon: Coalescent simulation of coding DNA sequences with recombination, migration and demography. BMC Bioinformatics 8: 458
- ^ Arenas, M. and Posada, D. (2010) Coalescent simulation of intracodon recombination. Genetics 184(2): 429–437
- ^ Browning, S.R. (2006) Multilocus association mapping using variable-length markov chains. American Journal of Human Genetics 78:903–913
- ^ Cornuet J.-M., Pudlo P., Veyssier J., Dehne-Garcia A., Gautier M., Leblois R., Marin J.-M., Estoup A. (2014) DIYABC v2.0: a software to make Approximate Bayesian Computation inferences about population history using Single Nucleotide Polymorphism, DNA sequence and microsatellite data. Bioinformatics '30': 1187–1189
- ^ Degnan, JH and LA Salter. 2005. Gene tree distributions under the coalescent process. Evolution 59(1): 24–37. pdf from coaltree.net/
- ^ Donnelly, P., Tavaré, S. (1995) Coalescents and genealogical structure under neutrality. Annual Review of Genetics 29:401–421
- ^ Ewing, G. and Hermisson J. (2010), MSMS: a coalescent simulation program including recombination, demographic structure and selection at a single locus, Bioinformatics 26:15
- ^ Hellenthal, G., Stephens M. (2006) msHOT: modifying Hudson's ms simulator to incorporate crossover and gene conversion hotspots Bioinformatics AOP
- ^ Hudson, Richard R. (1983a). "Testing the Constant-Rate Neutral Allele Model with Protein Sequence Data". Evolution 37 (1): 203–17. doi:10.2307/2408186. ISSN 1558-5646. JSTOR 2408186 – via JSTOR. (registration required (help)).
- ^ Hudson RR (1983b) Properties of a neutral allele model with intragenic recombination. Theoretical Population Biology 23:183–201.
- ^ Hudson RR (1991) Gene genealogies and the coalescent process. Oxford Surveys in Evolutionary Biology 7: 1–44
- ^ Hudson RR (2002) Generating samples under a Wright–Fisher neutral model. Bioinformatics 18:337–338
- ^ Kendal WS (2003) An exponential dispersion model for the distribution of human single nucleotide polymorphisms. Mol Biol Evol 20: 579–590
- Hein, J., Schierup, M., Wiuf C. (2004) Gene Genealogies, Variation and Evolution: A Primer in Coalescent Theory Oxford University Press ISBN 978-0-19-852996-5
- ^ Kaplan, N.L., Darden, T., Hudson, R.R. (1988) The coalescent process in models with selection. Genetics 120:819–829
- ^ Kingman, J. F. C. (1982). "On the Genealogy of Large Populations". Journal of Applied Probability 19: 27–43. doi:10.2307/3213548. ISSN 0021-9002. JSTOR 3213548 – via JSTOR. (registration required (help)).
- ^ Kingman, J.F.C. (2000) Origins of the coalescent 1974–1982. Genetics 156:1461–1463
- ^ Leblois R., Estoup A. and Rousset F. (2009) IBDSim: a computer program to simulate genotypic data under isolation by distance Molecular Ecology Resources 9:107–109
- ^ Liang L., Zöllner S., Abecasis G.R. (2007) GENOME: a rapid coalescent-based whole genome simulator. Bioinformatics 23: 1565–1567
- ^ Mailund, T., Schierup, M.H., Pedersen, C.N.S., Mechlenborg, P. J. M., Madsen, J.N., Schauser, L. (2005) CoaSim: A Flexible Environment for Simulating Genetic Data under Coalescent Models BMC Bioinformatics 6:252
- ^ Möhle, M., Sagitov, S. (2001) A classification of coalescent processes for haploid exchangeable population models The Annals of Probability 29:1547–1562
- ^ Morris, A. P., Whittaker, J. C., Balding, D. J. (2002) Fine-scale mapping of disease loci via shattered coalescent modeling of genealogies American Journal of Human Genetics 70:686–707
- ^ Neuhauser, C., Krone, S.M. (1997) The genealogy of samples in models with selection Genetics 145 519–534
- ^ Pitman, J. (1999) Coalescents with multiple collisions The Annals of Probability 27:1870–1902
- ^ Harding, Rosalind, M. 1998. New phylogenies: an introductory look at the coalescent. pp. 15–22, in Harvey, P. H., Brown, A. J. L., Smith, J. M., Nee, S. New uses for new phylogenies. Oxford University Press (ISBN 0198549849)
- ^ Rosenberg, N.A., Nordborg, M. (2002) Genealogical Trees, Coalescent Theory and the Analysis of Genetic Polymorphisms. Nature Reviews Genetics 3:380–390
- ^ Sagitov, S. (1999) The general coalescent with asynchronous mergers of ancestral lines Journal of Applied Probability 36:1116–1125
- ^ Schweinsberg, J. (2000) Coalescents with simultaneous multiple collisions Electronic Journal of Probability 5:1–50
- ^ Slatkin, M. (2001) Simulating genealogies of selected alleles in populations of variable size Genetic Research 145:519–534
- ^ Tajima, F. (1983) Evolutionary Relationship of DNA Sequences in finite populations. Genetics 105:437–460
- ^ Tavare S, Balding DJ, Griffiths RC & Donnelly P. 1997. Inferring coalescent times from DNA sequence data. Genetics 145: 505–518.
- ^ The international SNP map working group. 2001. A map of human genome variation containing 1.42 million single nucleotide polymorphisms. Nature 409: 928–933.
- ^ Zöllner S. and Pritchard J.K. (2005) Coalescent-Based Association Mapping and Fine Mapping of Complex Trait Loci Genetics 169:1071–1092
- ^ Rousset F. and Leblois R. (2007) Likelihood and Approximate Likelihood Analyses of Genetic Structure in a Linear Habitat: Performance and Robustness to Model Mis-Specification Molecular Biology and Evolution 24:2730–2745
Books
- Hein, J; Schierup, M. H., and Wiuf, C. Gene Genealogies, Variation and Evolution – A Primer in Coalescent Theory. Oxford University Press, 2005. ISBN 0-19-852996-1.
- Nordborg, M. (2001) Introduction to Coalescent Theory
- Chapter 7 in Balding, D., Bishop, M., Cannings, C., editors, Handbook of Statistical Genetics. Wiley ISBN 978-0-471-86094-5
- Wakeley J. (2006) An Introduction to Coalescent Theory Roberts & Co ISBN 0-9747077-5-9 Accompanying website with sample chapters
- ^ Rice SH. (2004). Evolutionary Theory: Mathematical and Conceptual Foundations. Sinauer Associates: Sunderland, MA. See esp. ch. 3 for detailed derivations.
- Berestycki N. "Recent progress in coalescent theory" 2009 ENSAIOS Matematicos vol.16
- Bertoin J. "Random Fragmentation and Coagulation Processes"., 2006. Cambridge Studies in Advanced Mathematics, 102. Cambridge University Press, Cambridge, 2006. ISBN 978-0-521-86728-3;
- Pitman J. "Combinatorial stochastic processes" Springer (2003)
External links
- EvoMath 3: Genetic Drift and Coalescence, Briefly — overview, with probability equations for genetic drift, and simulation graphs
| ||||||||||||||||||||||||||||||