Glutamate (neurotransmitter)
| Glutamate | |
|---|---|
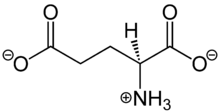 | |
| IUPAC name | 2-Aminopentanedioic acid |
| Synonyms | L-Glutamate |
| Abbreviation | GLU |
| Sources | almost every part of the nervous system |
| Targets | system-wide |
| Receptors | NMDA, AMPA, kainate, metabotropic |
| Agonists | NMDA, AMPA, kainic acid |
| Antagonists | AP5, ketamine, CNQX, kynurenic acid |
| Precursor | mainly dietary sources |
| Metabolizing enzyme | glutamate dehydrogenase |
| Database links | |
| CAS Registry Number |
56-86-0 |
| PubChem | CID: 104813 |
| ChemSpider |
591 |
| KEGG |
D0434 |
Glutamate is an amino acid, one of the twenty amino acids used to construct proteins, and as a consequence is found in high concentration in every part of the body. In the nervous system it plays a special additional role as a neurotransmitter: a chemical that nerve cells use to send signals to other cells. In fact glutamate is by a wide margin the most abundant neurotransmitter in the vertebrate nervous system.[1] It is used by every major excitatory information-transmitting pathway in the vertebrate brain, accounting in total for well over 90% of the synaptic connections in the human brain.
Chemical receptors for glutamate fall into three major classes, known as AMPA receptors, NMDA receptors, and metabotropic glutamate receptors. Many synapses use multiple types of glutamate receptors. AMPA receptors are ionotropic receptors specialized for fast excitation: in many synapses they produce excitatory electrical responses in their targets a fraction of a millisecond after being stimulated. NMDA receptors are also ionotropic, but they differ from AMPA receptors in being permeable, when activated, to calcium. Their properties make them particularly important for learning and memory. Metabotropic receptors act through second messenger systems to create slow, sustained effects on their targets. A fourth class, known as kainate receptors, are similar in many respects to AMPA receptors, but much less abundant.
Because of its role in synaptic plasticity, glutamate is involved in cognitive functions such as learning and memory in the brain.[2] The form of plasticity known as long-term potentiation takes place at glutamatergic synapses in the hippocampus, neocortex, and other parts of the brain.
Glutamate works not only as a point-to-point transmitter, but also through spill-over synaptic crosstalk between synapses in which summation of glutamate released from a neighboring synapse creates extrasynaptic signaling/volume transmission.[3] In addition, glutamate plays important roles in the regulation of growth cones and synaptogenesis during brain development as originally described by Mark Mattson.
Biosynthesis
Glutamate is a major constituent of a wide variety of proteins; consequently it is one of the most abundant amino acids in the human body.[1] Under ordinary conditions enough is obtained from the diet that there is no need for any to be synthesized. Nevertheless glutamate is formally classified as a non-essential amino acid, because it can be synthesized from alpha-Ketoglutaric acid, which is produced as part of the citric acid cycle by a series of reactions whose starting point is citrate. Glutamate cannot cross the blood-brain barrier unassisted, but it is actively transported into the nervous system by a high affinity transport system, which maintains its concentration in brain fluids at a fairly constant level.[4]
Glutamate itself serves as metabolic precursor for the neurotransmitter GABA, via the action of the enzyme glutamate decarboxylase.
Cellular effects
| Family | Type | Mechanism |
|---|---|---|
| AMPA | Ionotropic | Increase membrane permeability for sodium and potassium |
| kainate | Ionotropic | Increase membrane permeability for sodium and potassium |
| NMDA | Ionotropic, voltage gated | Increase membrane permeability for calcium |
| metabotropic Group I |
Gq-coupled | Increase IP3 and diacyl glycerol by activating phospholipase C |
| metabotropic Group II |
Gi/G0-coupled | Decrease intracellular levels of cAMP by inhibiting adenylate cyclase |
| metabotropic Group III |
Gi/G0-coupled | Decrease intracellular levels of cAMP by inhibiting adenylate cyclase |
Glutamate exerts its effects by binding to and activating cell surface receptors. In mammals, four families of glutamate receptors have been identified, known as AMPA receptors, kainate receptors, NMDA receptors, and metabotropic glutamate receptors. The first three families are ionotropic, meaning that when activated they open membrane channels that allow ions to pass through. The metabotropic family are G protein-coupled receptors, meaning that they exert their effects via a complex second messenger system.
Disease, disabilities, and pharmacology
Glutamate transporters,[5] EAAT and VGLUT, are found in neuronal and glial membranes. They rapidly remove glutamate from the extracellular space. In brain injury or disease, they often work in reverse, and excess glutamate can accumulate outside cells. This process causes calcium ions to enter cells via NMDA receptor channels, leading to neuronal damage and eventual cell death, and is called excitotoxicity. The mechanisms of cell death include
- Damage to mitochondria from excessively high intracellular Ca2+[6]
- Glu/Ca2+-mediated promotion of transcription factors for pro-apoptotic genes, or downregulation of transcription factors for anti-apoptotic genes
Excitotoxicity due to excessive glutamate release and impaired uptake occurs as part of the ischemic cascade and is associated with stroke,[7] autism,[8] some forms of intellectual disability, and diseases such as amyotrophic lateral sclerosis, lathyrism, and Alzheimer's disease.[7][9] In contrast, decreased glutamate release is observed under conditions of classical phenylketonuria[10] leading to developmental disruption of glutamate receptor expression.[11]
Glutamic acid has been implicated in epileptic seizures. Microinjection of glutamic acid into neurons produces spontaneous depolarisations around one second apart, and this firing pattern is similar to what is known as paroxysmal depolarizing shift in epileptic attacks. This change in the resting membrane potential at seizure foci could cause spontaneous opening of voltage-activated calcium channels, leading to glutamic acid release and further depolarization.
Comparative biology and evolution
Glutamate functions as a neurotransmitter in every type of animal that has a nervous system, including even ctenophores (comb jellies), which branched off from other phyla at a very early stage and lack any of the other neurotransmitters found ubiquitously among animals, such as serotonin and acetylcholine.[12] In fact, ctenophores have more distinct types of ionotropic glutamate receptors than any other known type of animal.[12] Activation of these receptors is capable of triggering muscle contraction, among other responses.[12]
Sponges do not have a nervous system, but even they apparently make use of glutamate for cell-to-cell signalling. Sponges possess glutamate receptors of the metabotropic type, and application of glutamate to a sponge can trigger a sort of whole-body "sneeze" response that sponges use to rid themselves of contaminants.[13] The genome of Trichoplax, a primitive blob-like organism that also has no nervous system, contains numerous glutamate receptors of the G-protein-coupled type, but their function is not known.[14]
In arthropods and nematodes, glutamate stimulates glutamate-gated chloride channels. The beta subunits of the receptor respond with very high affinity to glutamate and glycine.[15] Targeting these receptors has been the therapeutic goal of anthelmintic therapy using avermectins. Avermectins target the alpha subunit of glutamate-gated chloride channels with high affinity.[16] These receptors have also been described in arthropods, such as Drosophila melanogaster[17] and Lepeophtheirus salmonis.[18] Irreversible activation of these receptors with avermectins results in hyperpolarization at synapses and neuromuscular junctions resulting in flaccid paralysis and death of nematodes and arthropods.
History
The presence of glutamate in every part of the body as a building-block for protein made its special role in the nervous system difficult to recognize: its function as a neurotransmitter was not generally accepted until the 1970s, decades after the identification of acetylcholine, norepinephrine, and serotonin as neurotransmitters.[19] The first suggestion that glutamate might function as a transmitter came from T. Hayoshi in 1954, who was motivated by the finding that injections of glutamate into the cerebral ventricles of dogs could cause them to have seizures.[19] Other support for this idea soon appeared, but the majority of physiologists were skeptical, for a variety of theoretical and empirical reasons. Ironically, one of the most common reasons for skepticism was the universality of glutamate's excitatory effects in the CNS, which seemed inconsistent with the specificity expected of a neurotransmitter.[19] Other reasons for skepticism included a lack of known antagonists and the absence of a known mechanism for inactivation. A series of discoveries during the 1970s resolved most of these doubts, and by 1980 the compelling nature of the evidence was almost universally recognized.[19]
References
- 1 2 Meldrum, B. S. (2000). "Glutamate as a neurotransmitter in the brain: Review of physiology and pathology" (PDF). The Journal of nutrition 130 (4S Suppl): 1007S–1015S. PMID 10736372.
- ↑ McEntee, W. J.; Crook, T. H. (1993). "Glutamate: Its role in learning, memory, and the aging brain". Psychopharmacology 111 (4): 391–401. doi:10.1007/BF02253527. PMID 7870979.
- ↑ Okubo, Y.; Sekiya, H.; Namiki, S.; Sakamoto, H.; Iinuma, S.; Yamasaki, M.; Watanabe, M.; Hirose, K.; Iino, M. (2010). "Imaging extrasynaptic glutamate dynamics in the brain". Proceedings of the National Academy of Sciences 107 (14): 6526. doi:10.1073/pnas.0913154107.
- ↑ Smith QR (2000). "Transport of glutamate and other amino acids at the blood-brain barrier". J. Nutr. 130 (4S Suppl): 1016S–22S. PMID 10736373. Retrieved 3 December 2015.
- ↑ Shigeri, Yasushi; Seal, Rebecca P; Shimamoto, Keiko (2004). "Molecular pharmacology of glutamate transporters, EAATs and VGLUTs". Brain Research Reviews 45 (3): 250. doi:10.1016/j.brainresrev.2004.04.004. PMID 15210307.
- ↑ Manev, H.; Favaron, M.; Guidotti, A.; Costa, E. (1989). "Delayed increase of Ca2+ influx elicited by glutamate: Role in neuronal death". Molecular Pharmacology 36 (1): 106–112. PMID 2568579.
- 1 2 Robert Sapolsky (2005). "Biology and Human Behavior: The Neurological Origins of Individuality, 2nd edition". The Teaching Company.
see pages 19 and 20 of Guide Book
- ↑ Shinohe, Atsuko; Hashimoto, Kenji; Nakamura, Kazuhiko; Tsujii, Masatsugu; Iwata, Yasuhide; Tsuchiya, Kenji J.; Sekine, Yoshimoto; Suda, Shiro; Suzuki, Katsuaki (2006-12-30). "Increased serum levels of glutamate in adult patients with autism". Progress in Neuro-Psychopharmacology & Biological Psychiatry 30 (8): 1472–1477. doi:10.1016/j.pnpbp.2006.06.013. ISSN 0278-5846. PMID 16863675.
- ↑ Hynd, M; Scott, H. L.; Dodd, P. R. (2004). "Glutamate-mediated excitotoxicity and neurodegeneration in Alzheimer?s disease". Neurochemistry International 45 (5): 583–95. doi:10.1016/j.neuint.2004.03.007. PMID 15234100.
- ↑ Glushakov, AV; Dennis, DM; Sumners, C; Seubert, CN; Martynyuk, AE (Apr 1, 2003). "L-phenylalanine selectively depresses currents at glutamatergic excitatory synapses". Journal of neuroscience research 72 (1): 116–24. doi:10.1002/jnr.10569. PMID 12645085.
- ↑ Glushakov, AV; Glushakova, O; Varshney, M; Bajpai, LK; Sumners, C; Laipis, PJ; Embury, JE; Baker, SP; Otero, DH; Dennis, DM; Seubert, CN; Martynyuk, AE (February 2005). "Long-term changes in glutamatergic synaptic transmission in phenylketonuria". Brain : a journal of neurology 128 (Pt 2): 300–7. doi:10.1093/brain/awh354. PMID 15634735.
- 1 2 3 Moroz LL, Kocot KM, et al. (2014). "The ctenophore genome and the evolutionary origins of neural systems". Nature 510 (7503): 109–14. doi:10.1038/nature13400. PMC 4337882. PMID 24847885.
- ↑ Leys SP (2015). "Elements of a 'nervous system' in sponges". J. Exp. Biol. 218 (Pt 4): 581–91. doi:10.1242/jeb.110817. PMID 25696821.
- ↑ Krishnan A, Schiöth HB (2015). "The role of G protein-coupled receptors in the early evolution of neurotransmission and the nervous system". J. Exp. Biol. 218 (Pt 4): 562–71. doi:10.1242/jeb.110312. PMID 25696819.
- ↑ Laughton, D. L.; Wheeler, S. V.; Lunt, G. G.; Wolstenholme, A. J. (2002). "The β-Subunit of Caenorhabditis elegans Avermectin Receptor Responds to Glycine and is Encoded by Chromosome 1". Journal of Neurochemistry 64 (5): 2354–2357. doi:10.1046/j.1471-4159.1995.64052354.x. PMID 7536811.
- ↑ Cully, D. F.; Vassilatis, D. K.; Liu, K. K.; Paress, P. S.; Van Der Ploeg, L. H. T.; Schaeffer, J. M.; Arena, J. P. (1994). "Cloning of an avermectin-sensitive glutamate-gated chloride channel from Caenorhabditis elegans". Nature 371 (6499): 707–711. doi:10.1038/371707a0. PMID 7935817. horizontal tab character in
|title=at position 69 (help) - ↑ Cully, D.F., Paress, P.S., Liu, K.K., Schaeffer, J.M. and Arena, J.P. 1996. "Identification of a Drosophila melanogaster glutamate-gated chloride channel sensitive to the antiparasitic agent avermectin". J. Biol. Chem. '271, 20187-20191'
- ↑ Tribble, N. D.; Burka, J. F.; Kibenge, F. S. B. (2007). "Identification of the genes encoding for putative gamma aminobutyric acid (GABA) and glutamate-gated chloride channel (Glu Cl) alpha receptor subunits in sea lice (Lepeophtheirus salmonis)". Journal of Veterinary Pharmacology and Therapeutics 30 (2): 163. doi:10.1111/j.1365-2885.2007.00823.x. PMID 17348903.
- 1 2 3 4 Watkins JC (2000). "l-glutamate as a central neurotransmitter: looking back". Biochem. Soc. Trans. 28 (4): 297–309. PMID 10961913.