Antenna (biology)

Antennae (singular: antenna) in biology have historically been paired appendages used for sensing in arthropods. More recently, the term has also been applied to cilium structures present in most cell types of eukaryotes.
In arthropods, antennae are connected to the front-most segments. In crustaceans, they are biramous and present on the first two segments of the head, with the smaller pair known as antennules. All other arthropod groups – except chelicerates and proturans, which have none – have a single, uniramous pair of antennae. These antennae are jointed, at least at the base, and, in general, extend forward from the head. They are sensory organs, although the exact nature of what they sense and how they sense it is not the same in all groups, or always clear. Functions may variously include sensing touch, air motion, heat, vibration (sound), and especially olfaction (smell) or gustation (taste).
Insects

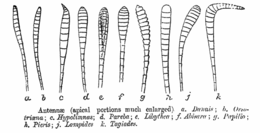
Antennae are the primary olfactory sensors of insects[1] and are accordingly well-equipped with a wide variety of sensilla (singular: sensillum). Paired, mobile, and segmented, they are located between the eyes on the forehead. Embryologically, they represent the appendages of the second head segment.[2]
All insects have antenna, however they may be greatly reduced in the larval forms. Amongst the non-insect classes of the Hexapoda, both Collembola and Diplura have antenna, but Protura do not.[3]
Structure

The three basic segments of the typical insect antenna are the scape or scapus (base), the pedicel or pedicellus (stem), and finally the flagellum, which often comprises many units known as flagellomeres.[4] The pedicel (the second segment) contains the Johnston's organ which is a collection of sensory cells.
The scape is mounted in a socket in a more or less ring-shaped sclerotised region called the torulus, often a raised portion of the insect's head capsule. The socket is closed off by the membrane into which the base of the scape is set. However, the antenna does not hang free on the membrane, but pivots on a rigidly sprung projection from the rim of the torulus. That projection on which the antenna pivots is called the antennifer. The whole structure enables the insect to move the antenna as a whole by applying internal muscles connected to the scape. The pedicel is flexibly connected to the distal end of the scape and its movements in turn can be controlled by muscular connections between the scape and pedicel. The number of flagellomeres can vary greatly between insect species, and often is of diagnostic importance.
True flagellomeres are connected by membranous linkage that permits movement, though the flagellum of "true" insects does not have any intrinsic muscles. Some other Arthropoda do however have intrinsic muscles throughout the flagellum. Such groups include the Symphyla, Collembola and Diplura. In many true insects, especially the more primitive groups such as Thysanura and Blattodea, the flagellum partly or entirely consists of a flexibly connected string of small ring-shaped annuli. The annuli are not true flagellomeres, and in a given insect species the number of annuli generally is not as consistent as the number of flagellomeres in most species.[4]
In many beetles and in the chalcidoid wasps, the apical flagellomeres form a club shape, and the collective term for the segments between the club and the antennal base is the funicle; traditionally in describing beetle anatomy, the term "funicle" refers to the segments between the club and the scape. However, traditionally in working on wasps the funicle is taken to comprise the segments between the club and the pedicel.[4]
Quite commonly the funicle beyond the pedicel is quite complex in Endopterygota such as beetles, moths and Hymenoptera, and one common adaptation is the ability to fold the antenna in the middle, at the joint between the pedicel and the flagellum. This gives an effect like a "knee bend", and such an antenna is said to be geniculate. Geniculate antennae are common in the Coleoptera and Hymenoptera. They are important for insects like ants that follow scent trails, for bees and wasps that need to "sniff" the flowers that they visit, and for beetles such as Scarabaeidae and Curculionidae that need to fold their antennae away when they self-protectively fold up all their limbs in defensive attitudes.
Because the funicle is without intrinsic muscles, it generally must move as a unit, in spite of being articulated. However, some funicles are complex and very mobile. For example, the Scarabaeidae have lamellate antennae that can be folded tightly for safety or spread openly for detecting odours or pheromones. The insect manages such actions by changes in blood pressure, by which it exploits elasticity in walls and membranes in the funicles, which are in effect erectile.[5]
In the groups with more uniform antennae (for example: millipedes), all segments are called antennomeres. Some groups have a simple or variously modified apical or subapical bristle called an arista (this may be especially well-developed in various Diptera).[6]
Functions
Olfactory receptors on the antennae bind to free-floating molecules, such as water vapour, and odours including pheromones. The neurons that possess these receptors signal this binding by sending action potentials down their axons to the antennal lobe in the brain. From there, neurons in the antennal lobes connect to mushroom bodies that identify the odour. The sum of the electrical potentials of the antennae to a given odour can be measured using an electroantennogram.[7]
In the case of the monarch butterfly, antennae are necessary for proper time-compensated solar compass orientation during migration, antennal clocks exist in monarchs, and they are likely to provide the primary timing mechanism for sun compass orientation.[8]
Crustaceans
Crustaceans bear two pairs of antennae. The first pair are uniramous and are often referred to as antennules, while the second pair are biramous, meaning that each antenna is composed of two parts, joined at their base .[9] In most adults, the antenna are sensory organs, but they are used by the nauplius larva for swimming. In some groups of crustaceans, such as the spiny lobsters and slipper lobsters, the second antennae are enlarged, while, in others, such as crabs, the antennae are reduced in size.




Cellular antennae

Within the biological and medical disciplines, recent discoveries have noted that primary cilia in many types of cells within eukaryotes serve as cellular antennae. These cilia play important roles in chemosensation, mechanosensation, and thermosensation. The current scientific understanding of primary cilia organelles views them as "sensory cellular antennae that coordinate a large number of cellular signaling pathways, sometimes coupling the signaling to ciliary motility or alternatively to cell division and differentiation."[10]
"Almost every vertebrate cell has a specialized cell surface projection called a primary cilium. …primary cilia are key participants in intercellular signaling. This new appreciation of primary cilia as cellular antennae that sense a wide variety of signals could help explain why ciliary defects underlie such a wide range of human disorders, including retinal degeneration, polycystic kidney disease, Bardet-Biedl syndrome, and neural tube defects."[11]
References
- ↑ Darby, Gene (1958). What is a butterfly?. Chicago: Benefic Press. p. 8. OCLC 1391997.
- ↑ Gullan, Penny J.; Cranston, Peter S. (2005). The Insects: an Outline of Entomology (3rd ed.). Oxford, UK: Blackwell Publishing. p. 38. ISBN 1-4051-1113-5.
- ↑ Chapman, Reginald Frederick (1998). The Insects: Structure and Function (4th ed.). New York: Cambridge University Press. p. 8. ISBN 0-521-57890-6.
- 1 2 3 Thomas A. Keil (1999). "Morphology and development of the peripheral olfactory organs". In Hansson, Bill S. Insect Olfaction (1st ed.). Springer. pp. 5–48. ISBN 978-3-540-65034-8.
- ↑ Pass, Guenther. The Anatomy and Ultrastructure of the Antennal Circulatory Organs in the Cockchafer Beetle Melolontha melolontha L. (Coleoptera, Scarabaeidae) Zoomorphology 01/1980; 96(1):77-89. DOI: 10.1007/BF00310078
- ↑ Lawrence, Eleanor, ed. (2005). Henderson's Dictionary of Biological Terms (13th ed.). Pearson Education. p. 51. ISBN 978-0-13-127384-9.
- ↑ "Electroantennography (EAG)". Georg-August-Universität Göttingen. Retrieved March 27, 2010.
- ↑ Merlin, Christine; Gegear, Robert J.; Reppert, Steven M. (September 2009). "Antennal circadian clocks coordinate sun compass orientation in migratory monarch butterflies". Science 325 (5948): 1700–1704. doi:10.1126/science.1176221. PMC 2754321. PMID 19779201.
- ↑ "Superphylum Arthropoda". University of Arizona.
- ↑ Satir, Peter; Christensen, Søren T. (June 2008). "Structure and function of mammalian cilia". Histochemistry and Cell Biology 129 (6): 687–693. doi:10.1007/s00418-008-0416-9. PMC 2386530. PMID 18365235.
- ↑ Singla, Veena; Reiter, Jeremy F. (August 2006). "The primary cilium as the cell's antenna: signaling at a sensory organelle". Science 313 (5787): 629–633. doi:10.1126/science.1124534. PMID 16888132.