Eyespot (mimicry)

An eyespot (sometimes ocellus) is an eye-like marking. They are found on butterflies, reptiles, felids, birds and fish.
Eyespots may be a form of mimicry in which a spot on the body of an animal resembles an eye of a different animal to deceive potential predator or prey species; to draw a predator's attention away from the most vulnerable body parts; or to appear as an inedible or even dangerous animal.[1] In larger animals, eyespots may play a role in intraspecies communication or courtship – the most well-known example is probably the eyespots on a peacock's display feathers.
The eye-like markings in some butterflies and moths, like the Bicyclus anynana, and certain other insects, as well as the Sunbittern (a bird) do not seem to serve only a mimicry function. In some other cases, the evolutionary function of such spots is also not understood. There is evidence that eyespots in butterflies are antipredator adaptations, either in deimatic displays to distract, startle or scare off predators, or to deflect attacks away from vital body parts.[2][3] Butterfly eyespots may also play a role in mate recognition and sexual selection,[4] playing a similar role to the eyespots found on larger organisms mentioned above.
Pattern formation of concentric spots is probably less complex than the conspicuous eyespots might suggest; eyespots may simply be morphogenetic "spandrels", not necessarily adaptations.
Evolution of the eyespot at the molecular level

Butterfly eyespots are formed as a result of a morphogenetic signaling center, or organizer called the focus. This signaling center induces its neighboring cells to produce specific pigments which pattern the eyespot.[5][6][7] Foci morphology and evolution appears to be the result of an altered regulatory circuit composed of the same patterning mechanisms conserved in insect wing patterning. This altered regulatory circuit is able to pattern both the anterior and posterior eyespots independent of the anterior/posterior wing compartmentalization restrictions.[7]
Breaking the highly conserved compartmentalization restrictions put forth by insect wing patterning, by redeployment of early developmental signaling sources like the canonical hedgehog (Hh) pathway, Distalless (Dll), and engrailed (en), this altered regulatory circuit is able to pattern both the anterior and posterior eyespots independent of the A/P compartmentalization restrictions through increased localized levels of hh signaling.[7][6][5] Increased hh signaling raises expression of its receptor ptc and transcription factor.[7] Normally, in the fruit-fly Drosophila, engrailed(en) acts in the posterior compartment to restrict ptc and ci expression to the anterior compartment by repressing transcription of ci, thereby preventing expression of ptc. However, This rogue expression of ptc and ci seen in the posterior compartment of some butterfly wing disks, is an indication that during the evolution of eyespots, there was a divergence in the repression or expression of these genes from that of drosophila.[6] From an evo-devo perspective, understanding the redeployment and plasticity of already existing regulatory mechanisms in butterfly eyespot loci development may give more insight into a fundamental mechanism for the evolution of novelty structures.[6][5]
Morphogenetic models
Eyespot patterning is thought to occur by morphogenetic activity, however the exact model and modes of action have yet to be elucidated. A number of experiments have been done to understand the mechanism of eyespot patterning all of which indicate morphogenetic activity.
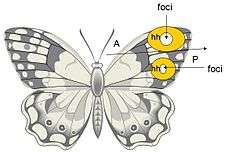
Effects of cautery on the butterfly wing eyespot foci are indicative of a long range signaling mechanism or morphogen gradient, both in its spacial and temporal aspects.[7] Cautery studies have shown that different results in eyespot size and color can result depending on the time in development at with the cauterizing damage is administered. While these findings support morphogenetic activity, they cannot simply be explained by a source/diffusion model.[7] Observations of cautery have given rise to two different explanations of how eyespots are patterned: 1) A source/threshold model, in which the focus is the morphogen source and, 2) the sink model, in which the focus is generating a gradient by removal of the morphogen at the foci.[7]
Further investigation of genes involved in wing development and morphogenetic activity has led to the discovery that hh has a primary role in the morphogenetic signaling center of the foci.[6] In a manner that is similar to Drosophila, hedgehog (Hh) is expressed in all cells in the posterior compartment of the developing butterfly wing during the mid fifth instar of butterfly wing development. However, in butterflies hh expression is significantly higher in those cells that flank the potential foci.[6] Higher transcription levels of hh along with other known associates of the hh pathway; patched (ptc) the hh receptor, and cubitus interruptus (ci) the hh transcription factor are seen throughout the mid to late fifth instar as well, which further implies a role for hedgehog signaling in eyespot development and patterning.[6] Furthermore, cells that are flanked by the cells expressing the highest level of hh signaling, are fated to become the foci, indicating that foci cell fate determination relies on high concentrations of hh in surrounding cells.[6]
Eyespots (ocelli) in felids
In members of the family Felidae (such as the leopard cat and leopard), the white circular markings on the backs of the ears are called ocelli, and they are functionally similar to eyespots in other animals. These ocelli on the back of felid ears serve social functions, such as signaling to kittens, and communicating the cat's mental state (excited, calm, aggressive, etc.) to conspecifics in the gloom of dense forest or in tall grass.
Eyespots in fish


Some fish species also display eyespots. The foureye butterflyfish gets its name from a large and conspicuous eyespot on each side of the body near the tail. A black vertical bar on the head runs through the true eye, making it hard to see.[9] This may deceive predators into attacking the tail rather than the more vulnerable head, and about the fish's likely direction of travel.[10] For the same reason, many juvenile fish display eyespots that may disappear during their adult phase.[11]
See also
| Wikimedia Commons has media related to Eyespot. |
References
- ↑ Stevens, Martin (2005). "The role of eyespots as anti-predator mechanisms, principally demonstrated in the Lepidoptera". Biological Reviews 80 (4): 573–588. doi:10.1017/S1464793105006810. PMID 16221330.
- ↑ Vallin, A., S. Jakobsson, J. Lind, and C. Wiklund (2005). "Prey survival by predator intimidation: an experimental study of peacock butterfly defence against blue tits". Proceedings Royal Society: Biological Sciences 272 (1569): 1203–1207. doi:10.1098/rspb.2004.3034.
- ↑ http://www.sify.com/news/female-butterflies-chase-males-when-it-s-cool-news-international-lbhpahdfgff.html
- ↑ Costanzo, K., and A. Monteiro (2006). "The use of chemical and visual cues in female choice in the butterfly Bicyclus anynana". Proceedings Royal Society: Biological Sciences 274: 845–851. doi:10.1098/rspb.2006.3729.
- 1 2 3 Breakfield, MP, Gates, J, Keys, D, Kesbeke, F, Wijngaarden, JP, Monteiro, A, French, V, Carroll SB. (November 1996). "Development, plasticity and evolution of butterfly eyespot patterns". Nature 384 (6606): 236–242. doi:10.1038/384236a0. PMID 12809139.
- 1 2 3 4 5 6 7 8 Keys, D. N, Lewis, D.L, Selegue, J. E, Pearson, BJ, Goodrich, LV, Johnson RL, Gates, J, Scott, MP, Carroll, SB (January 1999). "Recruitment of a hedgehog Regulatory Circuit in Butterfly Eyespot Evolution". Science 283 (5401): 532–534. doi:10.1126/science.283.5401.532. PMID 9915699.
- 1 2 3 4 5 6 7 French, V, Breakfield, PM. (September 1992). "The development of eyespot patterns on butterfly wings: morphogen sources or sinks?". Development 116: 103–109.
- ↑ Purple Flying Gurnard, Dactyloptena orientalis (Cuvier, 1829) Australian Museum. Updated: 15 September 2012. Retrieved: 2 November 2012.
- ↑ FishBaseFroese, Rainer and Pauly, Daniel, eds. (2009). "Chaetodon capistratus" in FishBase. July 2009 version.
- ↑ Cott, Hugh (1940). Adaptive Coloration in Animals. Oxford University Press. p. 373.
- ↑ Bos, A.R. (March 2015). "Soft corals provide microhabitat for camouflaged juveniles of the Blackspotted wrasse Macropharyngodon meleagris (Labridae)". Marine Biodiversity. doi:10.1007/s12526-015-0332-x.
Martin STEVENS, Graeme D RUXTON (2013) "Do animal eyespots really mimic eyes?" Current Zoology. Open Access article: http://www.currentzoology.org/site_media/onlinefirst/downloadable_file/2013/11/22/Martin_Stevens.pdf

| ||||||||||||||||||||||||
.jpg)