Evidence of common descent
| Part of a series on |
| Evolutionary biology |
|---|
 |
|
History of evolutionary theory |
|
Fields and applications
|
|
Evidence of common descent of living organisms has been discovered by scientists researching in a variety of disciplines over many decades and has demonstrated common descent of all life on Earth developing from a last universal ancestor. This evidence constructs the theoretical framework on which evolutionary theory rests, demonstrates that evolution does occur, and is able to show the natural processes that led to the emergence of Earth's biodiversity. Additionally, this evidence supports the modern evolutionary synthesis—the current scientific theory that explains how and why life changes over time. Evolutionary biologists document evidence of common descent by making testable predictions, testing hypotheses, and developing theories that illustrate and describe its causes.
Comparison of the DNA genetic sequences of organisms has revealed that organisms that are phylogenetically close have a higher degree of DNA sequence similarity than organisms that are phylogenetically distant. Further evidence for common descent comes from genetic detritus such as pseudogenes, regions of DNA that are orthologous to a gene in a related organism, but are no longer active and appear to be undergoing a steady process of degeneration from cumulative mutations.
Fossils are important for estimating when various lineages developed in geologic time. As fossilization is an uncommon occurrence, usually requiring hard body parts and death near a site where sediments are being deposited, the fossil record only provides sparse and intermittent information about the evolution of life. Scientific evidence of organisms prior to the development of hard body parts such as shells, bones and teeth is especially scarce, but exists in the form of ancient microfossils, as well as impressions of various soft-bodied organisms. The comparative study of the anatomy of groups of animals shows structural features that are fundamentally similar or homologous, demonstrating phylogenetic and ancestral relationships with other organisms, most especially when compared with fossils of ancient extinct organisms. Vestigial structures and comparisons in embryonic development are largely a contributing factor in anatomical resemblance in concordance with common descent. Since metabolic processes do not leave fossils, research into the evolution of the basic cellular processes is done largely by comparison of existing organisms' physiology and biochemistry. Many lineages diverged at different stages of development, so it is possible to determine when certain metabolic processes appeared by comparing the traits of the descendants of a common ancestor. Universal biochemical organization and molecular variance patterns in all organisms also show a direct correlation with common descent.
Further evidence comes from the field of biogeography because evolution with common descent provides the best and most thorough explanation for a variety of facts concerning the geographical distribution of plants and animals across the world. This is especially obvious in the field of insular biogeography. Combined with the theory of plate tectonics common descent provides a way to combine facts about the current distribution of species with evidence from the fossil record to provide a logically consistent explanation of how the distribution of living organisms has changed over time.
The development and spread of antibiotic resistant bacteria—like the spread of pesticide resistant forms of plants and insects—provides evidence that, evolution due to natural selection, is an ongoing process in the natural world. Alongside this are observed instances of the separation of populations of species into sets of new species (speciation). Speciation has been observed directly and indirectly in the lab and in nature. Multiple forms of such have been described and documented as examples for individual modes of speciation. Furthermore, evidence of common descent extends from direct laboratory experimentation with the selective breeding of organisms—historically and currently—and other controlled experiments involving many of the topics in the article. This article summarizes the varying disciplines that provide the evidence for evolution and the common descent of all life on Earth, accompanied by numerous, specialized examples.
Evidence from comparative physiology and biochemistry
Genetics

One of the strongest evidences for common descent comes from the study of gene sequences. Comparative sequence analysis examines the relationship between the DNA sequences of different species,[1] producing several lines of evidence that confirm Darwin's original hypothesis of common descent. If the hypothesis of common descent is true, then species that share a common ancestor inherited that ancestor's DNA sequence, as well as mutations unique to that ancestor. More closely related species have a greater fraction of identical sequence and shared substitutions compared to more distantly related species.
The simplest and most powerful evidence is provided by phylogenetic reconstruction. Such reconstructions, especially when done using slowly evolving protein sequences, are often quite robust and can be used to reconstruct a great deal of the evolutionary history of modern organisms (and even in some instances of the evolutionary history of extinct organisms, such as the recovered gene sequences of mammoths or Neanderthals). These reconstructed phylogenies recapitulate the relationships established through morphological and biochemical studies. The most detailed reconstructions have been performed on the basis of the mitochondrial genomes shared by all eukaryotic organisms, which are short and easy to sequence; the broadest reconstructions have been performed either using the sequences of a few very ancient proteins or by using ribosomal RNA sequence .
Phylogenetic relationships also extend to a wide variety of nonfunctional sequence elements, including repeats, transposons, pseudogenes, and mutations in protein-coding sequences that do not result in changes in amino-acid sequence. While a minority of these elements might later be found to harbor function, in aggregate they demonstrate that identity must be the product of common descent rather than common function .
Universal biochemical organisation and molecular variance patterns
All known extant (surviving) organisms are based on the same biochemical processes: genetic information encoded as nucleic acid (DNA, or RNA for many viruses), transcribed into RNA, then translated into proteins (that is, polymers of amino acids) by highly conserved ribosomes. Perhaps most tellingly, the Genetic Code (the "translation table" between DNA and amino acids) is the same for almost every organism, meaning that a piece of DNA in a bacterium codes for the same amino acid as in a human cell. ATP is used as energy currency by all extant life. A deeper understanding of developmental biology shows that common morphology is, in fact, the product of shared genetic elements.[2] For example, although camera-like eyes are believed to have evolved independently on many separate occasions,[3] they share a common set of light-sensing proteins (opsins), suggesting a common point of origin for all sighted creatures.[4][5] Another noteworthy example is the familiar vertebrate body plan, whose structure is controlled by the homeobox (Hox) family of genes.
DNA sequencing
Comparison of the DNA sequences allows organisms to be grouped by sequence similarity, and the resulting phylogenetic trees are typically congruent with traditional taxonomy, and are often used to strengthen or correct taxonomic classifications. Sequence comparison is considered a measure robust enough to correct erroneous assumptions in the phylogenetic tree in instances where other evidence is scarce. For example, neutral human DNA sequences are approximately 1.2% divergent (based on substitutions) from those of their nearest genetic relative, the chimpanzee, 1.6% from gorillas, and 6.6% from baboons.[6][7] Genetic sequence evidence thus allows inference and quantification of genetic relatedness between humans and other apes.[8][9] The sequence of the 16S ribosomal RNA gene, a vital gene encoding a part of the ribosome, was used to find the broad phylogenetic relationships between all extant life. The analysis, originally done by Carl Woese, resulted in the three-domain system, arguing for two major splits in the early evolution of life. The first split led to modern Bacteria and the subsequent split led to modern Archaea and Eukaryotes.
Some DNA sequences are shared by very different organisms. It has been predicted by the theory of evolution that the differences in such DNA sequences between two organisms should roughly resemble both the biological difference between them according to their anatomy and the time that had passed since these two organisms have separated in the course of evolution, as seen in fossil evidence. The rate of accumulating such changes should be low for some sequences, namely those that code for critical RNA or proteins, and high for others that code for less critical RNA or proteins; but for every specific sequence, the rate of change should be roughly constant over time. These results have been experimentally confirmed. Two examples are DNA sequences coding for rRNA, which is highly conserved, and DNA sequences coding for fibrinopeptides (amino acid chains that are discarded during the formation of fibrin), which are highly non-conserved.[10]
Endogenous retroviruses
Endogenous retroviruses (or ERVs) are remnant sequences in the genome left from ancient viral infections in an organism. The retroviruses (or virogenes) are always passed on to the next generation of that organism that received the infection. This leaves the virogene left in the genome. Because this event is rare and random, finding identical chromosomal positions of a virogene in two different species suggests common ancestry.[11] Cats (Felidae) present an notable instance of virogene sequences demonstrating common descent. The standard phylogenetic tree for Felidae have smaller cats (Felis chaus, Felis silvestris, Felis nigripes, and Felis catus) diverging from larger cats such as the subfamily Pantherinae and other carnivores. The fact that small cats have an ERV where the larger cats do not suggests that the gene was inserted into the ancestor of the small cats after the larger cats had diverged.[12] Another example of this is with humans and chimps. Humans contain numerous ERVs that comprise a considerable percentage of the genome. Sources vary, however, 1%[13] to 8%[14] has been proposed. Humans and chimps share seven different occurrences of virogenes while all primates share similar retroviruses congruent with phylogeny.[15]
Proteins
The proteomic evidence also supports the universal ancestry of life. Vital proteins, such as the ribosome, DNA polymerase, and RNA polymerase, are found in everything from the most primitive bacteria to the most complex mammals. The core part of the protein is conserved across all lineages of life, serving similar functions. Higher organisms have evolved additional protein subunits, largely affecting the regulation and protein-protein interaction of the core. Other overarching similarities between all lineages of extant organisms, such as DNA, RNA, amino acids, and the lipid bilayer, give support to the theory of common descent. Phylogenetic analyses of protein sequences from various organisms produce similar trees of relationship between all organisms.[16] The chirality of DNA, RNA, and amino acids is conserved across all known life. As there is no functional advantage to right- or left-handed molecular chirality, the simplest hypothesis is that the choice was made randomly by early organisms and passed on to all extant life through common descent. Further evidence for reconstructing ancestral lineages comes from junk DNA such as pseudogenes, "dead" genes that steadily accumulate mutations.[17]
Pseudogenes
Pseudogenes, also known as noncoding DNA, are extra DNA in a genome that do not get transcribed into RNA to synthesize proteins. Some of this noncoding DNA has known functions, but much of it has no known function and is called "Junk DNA". This is an example of a vestige since replicating these genes uses energy, making it a waste in many cases. A pseudogene can be produced when a coding gene accumulates mutations that prevent it from being transcribed, making it non-functional. But since it is not transcribed, it may disappear without affecting fitness, unless it has provided some beneficial function as non-coding DNA. Non-functional pseudogenes may be passed on to later species, thereby labeling the later species as descended from the earlier species.
Other mechanisms
There is also a large body of molecular evidence for a number of different mechanisms for large evolutionary changes, among them: genome and gene duplication, which facilitates rapid evolution by providing substantial quantities of genetic material under weak or no selective constraints; horizontal gene transfer, the process of transferring genetic material to another cell that is not an organism's offspring, allowing for species to acquire beneficial genes from each other; and recombination, capable of reassorting large numbers of different alleles and of establishing reproductive isolation. The Endosymbiotic theory explains the origin of mitochondria and plastids (e.g. chloroplasts), which are organelles of eukaryotic cells, as the incorporation of an ancient prokaryotic cell into ancient eukaryotic cell. Rather than evolving eukaryotic organelles slowly, this theory offers a mechanism for a sudden evolutionary leap by incorporating the genetic material and biochemical composition of a separate species. Evidence supporting this mechanism has been found in the protist Hatena: as a predator it engulfs a green algae cell, which subsequently behaves as an endosymbiont, nourishing Hatena, which in turn loses its feeding apparatus and behaves as an autotroph.[18][19]
Since metabolic processes do not leave fossils, research into the evolution of the basic cellular processes is done largely by comparison of existing organisms. Many lineages diverged when new metabolic processes appeared, and it is theoretically possible to determine when certain metabolic processes appeared by comparing the traits of the descendants of a common ancestor or by detecting their physical manifestations. As an example, the appearance of oxygen in the earth's atmosphere is linked to the evolution of photosynthesis.
Specific examples
Chromosome 2 in humans

Evidence for the evolution of Homo sapiens from a common ancestor with chimpanzees is found in the number of chromosomes in humans as compared to all other members of Hominidae. All hominidae have 24 pairs of chromosomes, except humans, who have only 23 pairs. Human chromosome 2 is a result of an end-to-end fusion of two ancestral chromosomes.[20][21]
The evidence for this includes:
- The correspondence of chromosome 2 to two ape chromosomes. The closest human relative, the common chimpanzee, has near-identical DNA sequences to human chromosome 2, but they are found in two separate chromosomes. The same is true of the more distant gorilla and orangutan.[22][23]
- The presence of a vestigial centromere. Normally a chromosome has just one centromere, but in chromosome 2 there are remnants of a second centromere.[24]
- The presence of vestigial telomeres. These are normally found only at the ends of a chromosome, but in chromosome 2 there are additional telomere sequences in the middle.[25]
Chromosome 2 thus presents very strong evidence in favour of the common descent of humans and other apes. According to J. W. IJdo, "We conclude that the locus cloned in cosmids c8.1 and c29B is the relic of an ancient telomere-telomere fusion and marks the point at which two ancestral ape chromosomes fused to give rise to human chromosome 2."[25]
Cytochrome c and b
A classic example of biochemical evidence for evolution is the variance of the ubiquitous (i.e. all living organisms have it, because it performs very basic life functions) protein Cytochrome c in living cells. The variance of cytochrome c of different organisms is measured in the number of differing amino acids, each differing amino acid being a result of a base pair substitution, a mutation. If each differing amino acid is assumed the result of one base pair substitution, it can be calculated how long ago the two species diverged by multiplying the number of base pair substitutions by the estimated time it takes for a substituted base pair of the cytochrome c gene to be successfully passed on. For example, if the average time it takes for a base pair of the cytochrome c gene to mutate is N years, the number of amino acids making up the cytochrome c protein in monkeys differ by one from that of humans, this leads to the conclusion that the two species diverged N years ago.
The primary structure of cytochrome c consists of a chain of about 100 amino acids. Many higher order organisms possess a chain of 104 amino acids.[26]
The cytochrome c molecule has been extensively studied for the glimpse it gives into evolutionary biology. Both chicken and turkeys have identical sequence homology (amino acid for amino acid), as do pigs, cows and sheep. Both humans and chimpanzees share the identical molecule, while rhesus monkeys share all but one of the amino acids:[27] the 66th amino acid is isoleucine in the former and threonine in the latter.[26]
What makes these homologous similarities particularly suggestive of common ancestry in the case of cytochrome c, in addition to the fact that the phylogenies derived from them match other phylogenies very well, is the high degree of functional redundancy of the cytochrome c molecule. The different existing configurations of amino acids do not significantly affect the functionality of the protein, which indicates that the base pair substitutions are not part of a directed design, but the result of random mutations that aren't subject to selection.[28]
In addition, Cytochrome b is commonly used as a region of mitochondrial DNA to determine phylogenetic relationships between organisms due to its sequence variability. It is considered most useful in determining relationships within families and genera. Comparative studies involving cytochrome b have resulted in new classification schemes and have been used to assign newly described species to a genus, as well as deepen the understanding of evolutionary relationships.[29]
Recent African origin of modern humans
Mathematical models of evolution, pioneered by the likes of Sewall Wright, Ronald Fisher and J. B. S. Haldane and extended via diffusion theory by Motoo Kimura, allow predictions about the genetic structure of evolving populations. Direct examination of the genetic structure of modern populations via DNA sequencing has allowed verification of many of these predictions. For example, the Out of Africa theory of human origins, which states that modern humans developed in Africa and a small sub-population migrated out (undergoing a population bottleneck), implies that modern populations should show the signatures of this migration pattern. Specifically, post-bottleneck populations (Europeans and Asians) should show lower overall genetic diversity and a more uniform distribution of allele frequencies compared to the African population. Both of these predictions are borne out by actual data from a number of studies.[30]
Evidence from comparative anatomy
Comparative study of the anatomy of groups of animals or plants reveals that certain structural features are basically similar. For example, the basic structure of all flowers consists of sepals, petals, stigma, style and ovary; yet the size, colour, number of parts and specific structure are different for each individual species. The neural anatomy of fossilized remains may also be compared using advanced imaging techniques.[31]
Atavisms
An atavism is an evolutionary throwback, such as traits reappearing that had disappeared generations ago.[32] Atavisms occur because genes for previously existing phenotypical features are often preserved in DNA, even though the genes are not expressed in some or most of the organisms possessing them.[33] Some examples of this are hind-legged snakes[34] or whales[35] (In July 1919 a humpback whale was caught by a ship operating out of Vancouver that had legs 4 ft 2 in (1.27 m) long.[36]); the extra toes of ungulates that do not even reach the ground,[37] chicken's teeth,[38] reemergence of sexual reproduction in Hieracium pilosella and Crotoniidae;[39] and humans with tails,[32] extra nipples, and large canine teeth.
Evolutionary developmental biology and embryonic development
Evolutionary developmental biology is the biological field that compares the developmental process of different organisms to determine ancestral relationships between species. A large variety of organism's genomes contain a small fraction of genes that control the organisms development. Hox genes are an example of these types of nearly universal genes in organisms pointing to an origin of common ancestry. Embryological evidence comes from the development of organisms at the embryological level with the comparison of different organisms embryos similarity. Remains of ancestral traits often appear and disappear in different stages of the embryological development process. Examples include such as hair growth and loss (lanugo) during human development;[40] development and degeneration of a yolk sac; terrestrial frogs and salamanders passing through the larval stage within the egg—with features of typically aquatic larvae—but hatch ready for life on land;[41] and the appearance of gill-like structures (pharyngeal arch) in vertebrate embryo development. Note that in fish, the arches continue to develop as branchial arches while in humans, for example, they give rise to a variety of structures within the head and neck.
Homologous structures and divergent (adaptive) evolution
If widely separated groups of organisms are originated from a common ancestry, they are expected to have certain basic features in common. The degree of resemblance between two organisms should indicate how closely related they are in evolution:
- Groups with little in common are assumed to have diverged from a common ancestor much earlier in geological history than groups with a lot in common;
- In deciding how closely related two animals are, a comparative anatomist looks for structures that are fundamentally similar, even though they may serve different functions in the adult. Such structures are described as homologous and suggest a common origin.
- In cases where the similar structures serve different functions in adults, it may be necessary to trace their origin and embryonic development. A similar developmental origin suggests they are the same structure, and thus likely derived from a common ancestor.
When a group of organisms share a homologous structure that is specialized to perform a variety of functions to adapt different environmental conditions and modes of life, it is called adaptive radiation. The gradual spreading of organisms with adaptive radiation is known as divergent evolution.
Nested hierarchies and classification
Taxonomy is based on the fact that all organisms are related to each other in nested hierarchies based on shared characteristics. Most existing species can be organized rather easily in a nested hierarchical classification. This is evident from the Linnaean classification scheme. Based on shared derived characters, closely related organisms can be placed in one group (such as a genus), several genera can be grouped together into one family, several families can be grouped together into an order, etc.[42] The existence of these nested hierarchies was recognized by many biologists before Darwin, but he showed that his theory of evolution with its branching pattern of common descent could explain them.[42][43] Darwin described how common descent could provide a logical basis for classification:[44]
| “ | All the foregoing rules and aids and difficulties in classification are explained, if I do not greatly deceive myself, on the view that the natural system is founded on descent with modification; that the characters which naturalists consider as showing true affinity between any two or more species, are those which have been inherited from a common parent, and, in so far, all true classification is genealogical; that community of descent is the hidden bond which naturalists have been unconsciously seeking, ... | ” | |
Evolutionary trees
An evolutionary tree (of Amniota, for example, the last common ancestor of mammals and reptiles, and all its descendants) illustrates the initial conditions causing evolutionary patterns of similarity (e.g., all Amniotes produce an egg that possesses the amnios) and the patterns of divergence amongst lineages (e.g., mammals and reptiles branching from the common ancestry in Amniota). Evolutionary trees provide conceptual models of evolving systems once thought limited in the domain of making predictions out of the theory.[45] However, the method of phylogenetic bracketing is used to infer predictions with far greater probability than raw speculation. For example, paleontologists use this technique to make predictions about nonpreservable traits in fossil organisms, such as feathered dinosaurs, and molecular biologists use the technique to posit predictions about RNA metabolism and protein functions.[46][47] Thus evolutionary trees are evolutionary hypotheses that refer to specific facts, such as the characteristics of organisms (e.g., scales, feathers, fur), providing evidence for the patterns of descent, and a causal explanation for modification (i.e., natural selection or neutral drift) in any given lineage (e.g., Amniota). Evolutionary biologists test evolutionary theory using phylogenetic systematic methods that measure how much the hypothesis (a particular branching pattern in an evolutionary tree) increases the likelihood of the evidence (the distribution of characters among lineages).[48][49][50] The severity of tests for a theory increases if the predictions "are the least probable of being observed if the causal event did not occur."[51] "Testability is a measure of how much the hypothesis increases the likelihood of the evidence."[52]
Vestigial structures
A strong and direct evidence for common descent comes from vestigial structures.[53] Rudimentary body parts, those that are smaller and simpler in structure than corresponding parts in the ancestral species, are called vestigial organs. They are usually degenerated or underdeveloped. The existence of vestigial organs can be explained in terms of changes in the environment or modes of life of the species. Those organs are typically functional in the ancestral species but are now either nonfunctional or re-purposed. Examples are the pelvic girdles of whales, haltere (hind wings) of flies and mosquitos, wings of flightless birds such as ostriches, and the leaves of some xerophytes (e.g. cactus) and parasitic plants (e.g. dodder). However, vestigial structures may have their original function replaced with another. For example, the halteres in dipterists help balance the insect while in flight and the wings of ostriches are used in mating rituals.
Specific examples
.png)

(A) Primitive state — biting and chewing: e.g. grasshopper. Strong mandibles and maxillae for manipulating food.
(B) Ticking and biting: e.g. honey bee. Labium long to lap up nectar; mandibles chew pollen and mould wax.
(C) Sucking: e.g. butterfly. Labrum reduced; mandibles lost; maxillae long forming sucking tube.


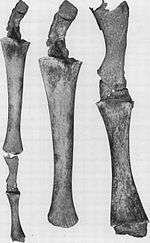
Hind structures in whales
Whales possess internally reduced hind parts such as the pelvis and hind legs (Fig. 5a).[54][55] Occasionally, the genes that code for longer extremities cause a modern whale to develop legs. On October 28, 2006, a four-finned bottlenose dolphin was caught and studied due to its extra set of hind limbs.[56] These legged Cetacea display an example of an atavism predicted from their common ancestry.
Insect mouthparts and appendages
Many different species of insects have mouthparts derived from the same embryonic structures, indicating that the mouthparts are modifications of a common ancestor's original features. These include a labrum (upper lip), a pair of mandibles, a hypopharynx (floor of mouth), a pair of maxillae, and a labium. (Fig. 5b) Evolution has caused enlargement and modification of these structures in some species, while it has caused the reduction and loss of them in other species. The modifications enable the insects to exploit a variety of food materials.
Insect mouthparts and antennae are considered homologues of insect legs. Parallel developments are seen in some arachnids: The anterior pair of legs may be modified as analogues of antennae, particularly in whip scorpions, which walk on six legs. These developments provide support for the theory that complex modifications often arise by duplication of components, with the duplicates modified in different directions.
Pelvic structure of dinosaurs
Similar to the pentadactyl limb in mammals, the earliest dinosaurs split into two distinct orders—the saurischia and ornithischia. They are classified as one or the other in accordance with what the fossils demonstrate. Figure 5c, shows that early saurischians resembled early ornithischians. The pattern of the pelvis in all species of dinosaurs is an example of homologous structures. Each order of dinosaur has slightly differing pelvis bones providing evidence of common descent. Additionally, modern birds show a similarity to ancient saurischian pelvic structures indicating the evolution of birds from dinosaurs. This can also be seen in Figure 5c as the Aves branch off the Theropoda suborder.
Pentadactyl limb
The pattern of limb bones called pentadactyl limb is an example of homologous structures (Fig. 5d). It is found in all classes of tetrapods (i.e. from amphibians to mammals). It can even be traced back to the fins of certain fossil fishes from which the first amphibians evolved such as tiktaalik. The limb has a single proximal bone (humerus), two distal bones (radius and ulna), a series of carpals (wrist bones), followed by five series of metacarpals (palm bones) and phalanges (digits). Throughout the tetrapods, the fundamental structures of pentadactyl limbs are the same, indicating that they originated from a common ancestor. But in the course of evolution, these fundamental structures have been modified. They have become superficially different and unrelated structures to serve different functions in adaptation to different environments and modes of life. This phenomenon is shown in the forelimbs of mammals. For example:
- In the monkey, the forelimbs are much elongated to form a grasping hand for climbing and swinging among trees.
- In the pig, the first digit is lost, and the second and fifth digits are reduced. The remaining two digits are longer and stouter than the rest and bear a hoof for supporting the body.
- In the horse, the forelimbs are adapted for support and running by great elongation of the third digit bearing a hoof.
- The mole has a pair of short, spade-like forelimbs for burrowing.
- The anteater uses its enlarged third digit for tearing down ant hills and termite nests.
- In the whale, the forelimbs become flippers for steering and maintaining equilibrium during swimming.
- In the bat, the forelimbs have turned into wings for flying by great elongation of four digits, while the hook-like first digit remains free for hanging from trees.
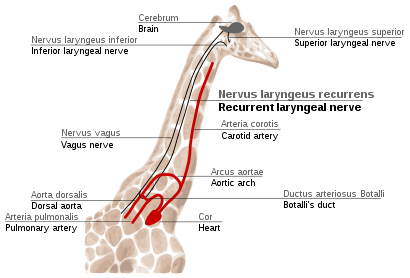
Recurrent laryngeal nerve in giraffes
The recurrent laryngeal nerve is a fourth branch of the vagus nerve, which is a cranial nerve. In mammals, its path is unusually long. As a part of the vagus nerve, it comes from the brain, passes through the neck down to heart, rounds the dorsal aorta and returns up to the larynx, again through the neck. (Fig. 5e)
This path is suboptimal even for humans, but for giraffes it becomes even more suboptimal. Due to the lengths of their necks, the recurrent laryngeal nerve may be up to 4m long (13 ft), despite its optimal route being a distance of just several inches.
The indirect route of this nerve is the result of evolution of mammals from fish, which had no neck and had a relatively short nerve that innervated one gill slit and passed near the gill arch. Since then, the gill it innervated has become the larynx and the gill arch has become the dorsal aorta in mammals.[57][58]
Route of the vas deferens

Similar to the laryngeal nerve in giraffes, the vas deferens is part of the male anatomy of many vertebrates; it transports sperm from the epididymis in anticipation of ejaculation. In humans, the vas deferens routes up from the testicle, looping over the ureter, and back down to the urethra and penis. It has been suggested that this is due to the descent of the testicles during the course of human evolution—likely associated with temperature. As the testicles descended, the vas deferens lengthened to accommodate the accidental "hook" over the ureter.[58][59]
Evidence from paleontology
.jpg)
When organisms die, they often decompose rapidly or are consumed by scavengers, leaving no permanent evidences of their existence. However, occasionally, some organisms are preserved. The remains or traces of organisms from a past geologic age embedded in rocks by natural processes are called fossils. They are extremely important for understanding the evolutionary history of life on Earth, as they provide direct evidence of evolution and detailed information on the ancestry of organisms. Paleontology is the study of past life based on fossil records and their relations to different geologic time periods.
For fossilization to take place, the traces and remains of organisms must be quickly buried so that weathering and decomposition do not occur. Skeletal structures or other hard parts of the organisms are the most commonly occurring form of fossilized remains (Paul, 1998), (Behrensmeyer, 1980) and (Martin, 1999). There are also some trace "fossils" showing moulds, cast or imprints of some previous organisms.
As an animal dies, the organic materials gradually decay, such that the bones become porous. If the animal is subsequently buried in mud, mineral salts infiltrate into the bones and gradually fill up the pores. The bones harden into stones and are preserved as fossils. This process is known as petrification. If dead animals are covered by wind-blown sand, and if the sand is subsequently turned into mud by heavy rain or floods, the same process of mineral infiltration may occur. Apart from petrification, the dead bodies of organisms may be well preserved in ice, in hardened resin of coniferous trees (amber), in tar, or in anaerobic, acidic peat. Fossilization can sometimes be a trace, an impression of a form. Examples include leaves and footprints, the fossils of which are made in layers that then harden.
Fossil record

It is possible to decipher how a particular group of organisms evolved by arranging its fossil record in a chronological sequence. Such a sequence can be determined because fossils are mainly found in sedimentary rock. Sedimentary rock is formed by layers of silt or mud on top of each other; thus, the resulting rock contains a series of horizontal layers, or strata. Each layer contains fossils typical for a specific time period when they formed. The lowest strata contain the oldest rock and the earliest fossils, while the highest strata contain the youngest rock and more recent fossils.
A succession of animals and plants can also be seen from fossil discoveries. By studying the number and complexity of different fossils at different stratigraphic levels, it has been shown that older fossil-bearing rocks contain fewer types of fossilized organisms, and they all have a simpler structure, whereas younger rocks contain a greater variety of fossils, often with increasingly complex structures.[60]
For many years, geologists could only roughly estimate the ages of various strata and the fossils found. They did so, for instance, by estimating the time for the formation of sedimentary rock layer by layer. Today, by measuring the proportions of radioactive and stable elements in a given rock, the ages of fossils can be more precisely dated by scientists. This technique is known as radiometric dating.
Throughout the fossil record, many species that appear at an early stratigraphic level disappear at a later level. This is interpreted in evolutionary terms as indicating the times when species originated and became extinct. Geographical regions and climatic conditions have varied throughout Earth's history. Since organisms are adapted to particular environments, the constantly changing conditions favoured species that adapted to new environments through the mechanism of natural selection.
Extent of the fossil record



Despite the relative rarity of suitable conditions for fossilization, approximately 250,000 fossil species are known.[62] The number of individual fossils this represents varies greatly from species to species, but many millions of fossils have been recovered: for instance, more than three million fossils from the last Ice Age have been recovered from the La Brea Tar Pits in Los Angeles.[63] Many more fossils are still in the ground, in various geological formations known to contain a high fossil density, allowing estimates of the total fossil content of the formation to be made. An example of this occurs in South Africa's Beaufort Formation (part of the Karoo Supergroup, which covers most of South Africa), which is rich in vertebrate fossils, including therapsids (reptile/mammal transitional forms).[64] It has been estimated that this formation contains 800 billion vertebrate fossils.[65]
Limitations
The fossil record is an important source for scientists when tracing the evolutionary history of organisms. However, because of limitations inherent in the record, there are not fine scales of intermediate forms between related groups of species. This lack of continuous fossils in the record is a major limitation in tracing the descent of biological groups. When transitional fossils are found that show intermediate forms in what had previously been a gap in knowledge, they are often popularly referred to as "missing links".
There is a gap of about 100 million years between the beginning of the Cambrian period and the end of the Ordovician period. The early Cambrian period was the period from which numerous fossils of sponges, cnidarians (e.g., jellyfish), echinoderms (e.g., eocrinoids), molluscs (e.g., snails) and arthropods (e.g., trilobites) are found. The first animal that possessed the typical features of vertebrates, the Arandaspis, was dated to have existed in the later Ordovician period. Thus few, if any, fossils of an intermediate type between invertebrates and vertebrates have been found, although likely candidates include the Burgess Shale animal, Pikaia gracilens,[66] and its Maotianshan shales relatives, Myllokunmingia, Yunnanozoon, Haikouella lanceolata,[67] and Haikouichthys.[68]
Some of the reasons for the incompleteness of fossil records are:
- In general, the probability that an organism becomes fossilized is very low;
- Some species or groups are less likely to become fossils because they are soft-bodied;
- Some species or groups are less likely to become fossils because they live (and die) in conditions that are not favourable for fossilization;
- Many fossils have been destroyed through erosion and tectonic movements;
- Most fossils are fragmentary;
- Some evolutionary change occurs in populations at the limits of a species' ecological range, and as these populations are likely small, the probability of fossilization is lower (see punctuated equilibrium);
- Similarly, when environmental conditions change, the population of a species is likely to be greatly reduced, such that any evolutionary change induced by these new conditions is less likely to be fossilized;
- Most fossils convey information about external form, but little about how the organism functioned;
- Using present-day biodiversity as a guide, this suggests that the fossils unearthed represent only a small fraction of the large number of species of organisms that lived in the past.
Specific examples
Evolution of the horse

Due to an almost-complete fossil record found in North American sedimentary deposits from the early Eocene to the present, the horse provides one of the best examples of evolutionary history (phylogeny).
This evolutionary sequence starts with a small animal called Hyracotherium (commonly referred to as Eohippus), which lived in North America about 54 million years ago then spread across to Europe and Asia. Fossil remains of Hyracotherium show it to have differed from the modern horse in three important respects: it was a small animal (the size of a fox), lightly built and adapted for running; the limbs were short and slender, and the feet elongated so that the digits were almost vertical, with four digits in the forelimbs and three digits in the hindlimbs; and the incisors were small, the molars having low crowns with rounded cusps covered in enamel.[69]
The probable course of development of horses from Hyracotherium to Equus (the modern horse) involved at least 12 genera and several hundred species. The major trends seen in the development of the horse to changing environmental conditions may be summarized as follows:
- Increase in size (from 0.4 m to 1.5 m — from 15in to 60in);
- Lengthening of limbs and feet;
- Reduction of lateral digits;
- Increase in length and thickness of the third digit;
- Increase in width of incisors;
- Replacement of premolars by molars; and
- Increases in tooth length, crown height of molars.
Fossilized plants found in different strata show that the marshy, wooded country in which Hyracotherium lived became gradually drier. Survival now depended on the head being in an elevated position for gaining a good view of the surrounding countryside, and on a high turn of speed for escape from predators, hence the increase in size and the replacement of the splayed-out foot by the hoofed foot. The drier, harder ground would make the original splayed-out foot unnecessary for support. The changes in the teeth can be explained by assuming that the diet changed from soft vegetation to grass. A dominant genus from each geological period has been selected to show the slow alteration of the horse lineage from its ancestral to its modern form.[70]
Transition from fish to amphibians
Prior to 2004, paleontologists had found fossils of amphibians with necks, ears, and four legs, in rock no older than 365 million years old. In rocks more than 385 million years old they could only find fish, without these amphibian characteristics. Evolutionary theory predicted that since amphibians evolved from fish, an intermediate form should be found in rock dated between 365 and 385 million years ago. Such an intermediate form should have many fish-like characteristics, conserved from 385 million years ago or more, but also have many amphibian characteristics as well. In 2004, an expedition to islands in the Canadian arctic searching specifically for this fossil form in rocks that were 375 million years old discovered fossils of Tiktaalik.[71] Some years later, however, scientists in Poland found evidence of fossilised tetrapod tracks predating Tiktaalik.[72]
Evidence from geographic distribution
Data about the presence or absence of species on various continents and islands (biogeography) can provide evidence of common descent and shed light on patterns of speciation.
Continental distribution
All organisms are adapted to their environment to a greater or lesser extent. If the abiotic and biotic factors within a habitat are capable of supporting a particular species in one geographic area, then one might assume that the same species would be found in a similar habitat in a similar geographic area, e.g. in Africa and South America. This is not the case. Plant and animal species are discontinuously distributed throughout the world:
- Africa has Old World monkeys, apes, elephants, leopards, giraffes, and hornbills.
- South America has New World monkeys, cougars, jaguars, sloths, llamas, and toucans.
- Deserts in North and South America have native cacti, but deserts in Africa, Asia, and Australia have succulent (apart from Rhipsalis baccifera)[73] which are native euphorbs that resemble cacti but are very different.
Even greater differences can be found if Australia is taken into consideration, though it occupies the same latitude as much of South America and Africa. Marsupials like kangaroos, bandicoots, and quolls make up about half of Australia's indigenous mammal species.[74] By contrast, marsupials are today totally absent from Africa and form a smaller portion of the mammalian fauna of South America, where opossums, shrew opossums, and the monito del monte occur. The only living representatives of primitive egg-laying mammals (monotremes) are the echidnas and the platypus. The short-beaked echidna (Tachyglossus aculeatus) and its subspecies populate Australia, Tasmania, New Guinea, and Kangaroo Island while the long-beaked echidna (Zaglossus bruijni) lives only in New Guinea. The platypus lives in the waters of eastern Australia. They have been introduced to Tasmania, King Island, and Kangaroo Island. These Monotremes are totally absent in the rest of the world.[75] On the other hand, Australia is missing many groups of placental mammals that are common on other continents (carnivorans, artiodactyls, shrews, squirrels, lagomorphs), although it does have indigenous bats and murine rodents; many other placentals, such as rabbits and foxes, have been introduced there by humans.
Other animal distribution examples include bears, located on all continents excluding Africa, Australia and Antarctica, and the polar bear only located solely in the Arctic Circle and adjacent land masses.[76] Penguins are located only around the South Pole despite similar weather conditions at the North Pole. Families of sirenians are distributed exclusively around the earth's waters, where manatees are located in western Africa waters, northern South American waters, and West Indian waters only while the related family, the dugongs, are located only in Oceanic waters north of Australia, and the coasts surrounding the Indian Ocean Additionally, the now extinct Steller's sea cow resided in the Bering Sea.[77]
The same kinds of fossils are found from areas known to be adjacent to one another in the past but that, through the process of continental drift, are now in widely divergent geographic locations. For example, fossils of the same types of ancient amphibians, arthropods and ferns are found in South America, Africa, India, Australia and Antarctica, which can be dated to the Paleozoic Era, when these regions were united as a single landmass called Gondwana.[78] Sometimes the descendants of these organisms can be identified and show unmistakable similarity to each other, even though they now inhabit very different regions and climates.
Island biogeography

Types of species found on islands
Evidence from island biogeography has played an important and historic role in the development of evolutionary biology. For purposes of biogeography, islands are divided into two classes. Continental islands are islands like Great Britain, and Japan that have at one time or another been part of a continent. Oceanic islands, like the Hawaiian islands, the Galápagos Islands and St. Helena, on the other hand are islands that have formed in the ocean and never been part of any continent. Oceanic islands have distributions of native plants and animals that are unbalanced in ways that make them distinct from the biotas found on continents or continental islands. Oceanic islands do not have native terrestrial mammals (they do sometimes have bats and seals), amphibians, or fresh water fish. In some cases they have terrestrial reptiles (such as the iguanas and giant tortoises of the Galápagos Islands) but often (for example Hawaii) they do not. This despite the fact that when species such as rats, goats, pigs, cats, mice, and cane toads, are introduced to such islands by humans they often thrive. Starting with Charles Darwin, many scientists have conducted experiments and made observations that have shown that the types of animals and plants found, and not found, on such islands are consistent with the theory that these islands were colonized accidentally by plants and animals that were able to reach them. Such accidental colonization could occur by air, such as plant seeds carried by migratory birds, or bats and insects being blown out over the sea by the wind, or by floating from a continent or other island by sea, as for example by some kinds of plant seeds like coconuts that can survive immersion in salt water, and reptiles that can survive for extended periods on rafts of vegetation carried to sea by storms.[79]
Endemism
Many of the species found on remote islands are endemic to a particular island or group of islands, meaning they are found nowhere else on earth. Examples of species endemic to islands include many flightless birds of New Zealand, lemurs of Madagascar, the Komodo dragon of Komodo,[80] the dragon's blood tree of Socotra,[81] Tuatara of New Zealand,[82][83] and others. However many such endemic species are related to species found on other nearby islands or continents; the relationship of the animals found on the Galápagos Islands to those found in South America is a well-known example.[79] All of these facts, the types of plants and animals found on oceanic islands, the large number of endemic species found on oceanic islands, and the relationship of such species to those living on the nearest continents, are most easily explained if the islands were colonized by species from nearby continents that evolved into the endemic species now found there.[79]
Other types of endemism do not have to include, in the strict sense, islands. Islands can mean isolated lakes or remote and isolated areas. Examples of these would include the highlands of Ethiopia, Lake Baikal, fynbos of South Africa, forests of New Caledonia, and others. Examples of endemic organisms living in isolated areas include the kagu of New Caledonia,[84] cloud rats of the Luzon tropical pine forests of the Philippines,[85][86] the boojum tree (Fouquieria columnaris) of the Baja California peninsula,[87] the Baikal seal[88] and the omul of Lake Baikal.
Adaptive radiations
Oceanic islands are frequently inhabited by clusters of closely related species that fill a variety of ecological niches, often niches that are filled by very different species on continents. Such clusters, like the finches of the Galápagos, Hawaiian honeycreepers, members of the sunflower family on the Juan Fernandez Archipelago and wood weevils on St. Helena are called adaptive radiations because they are best explained by a single species colonizing an island (or group of islands) and then diversifying to fill available ecological niches. Such radiations can be spectacular; 800 species of the fruit fly family Drosophila, nearly half the world's total, are endemic to the Hawaiian islands. Another illustrative example from Hawaii is the silversword alliance, which is a group of thirty species found only on those islands. Members range from the silverswords that flower spectacularly on high volcanic slopes to trees, shrubs, vines and mats that occur at various elevations from mountain top to sea level, and in Hawaiian habitats that vary from deserts to rainforests. Their closest relatives outside Hawaii, based on molecular studies, are tarweeds found on the west coast of North America. These tarweeds have sticky seeds that facilitate distribution by migrant birds.[89] Additionally, nearly all of the species on the island can be crossed and the hybrids are often fertile,[41] and they have been hybridized experimentally with two of the west coast tarweed species as well.[90] Continental islands have less distinct biota, but those that have been long separated from any continent also have endemic species and adaptive radiations, such as the 75 lemur species of Madagascar, and the eleven extinct moa species of New Zealand.[79][91]
Ring species

In biology, a ring species is a connected series of neighboring populations that can interbreed with relatively closely related populations, but for which there exist at least two "end" populations in the series that are too distantly related to interbreed. Often such non-breeding-though-genetically-connected populations co-exist in the same region thus creating a "ring". Ring species provide important evidence of evolution in that they illustrate what happens over time as populations genetically diverge, and are special because they represent in living populations what normally happens over time between long deceased ancestor populations and living populations. If any of the populations intermediate between the two ends of the ring were gone they would not be a continuous line of reproduction and each side would be a different species.[92][93]
Specific examples



Distribution of Glossopteris
The combination of continental drift and evolution can sometimes be used to predict what will be found in the fossil record. Glossopteris is an extinct species of seed fern plants from the Permian. Glossopteris appears in the fossil record around the beginning of the Permian on the ancient continent of Gondwana.[94] Continental drift explains the current biogeography of the tree. Present day Glossopteris fossils are found in Permian strata in southeast South America, southeast Africa, all of Madagascar, northern India, all of Australia, all of New Zealand, and scattered on the southern and northern edges of Antarctica. During the Permian, these continents were connected as Gondwana (see figure 6a) in agreement with magnetic striping, other fossil distributions, and glacial scratches pointing away from the temperate climate of the South Pole during the Permian.[95][96]
Metatherian Distribution
The history of metatherians (the clade containing marsupials and their extinct, primitive ancestors) provides an example of how evolutionary theory and the movement of continents can be combined to make predictions concerning fossil stratigraphy and distribution. The oldest metatherian fossils are found in present-day China.[97] Metatherians spread westward into modern North America (still attached to Eurasia) and then to South America, which was connected to North America until around 65 mya. Marsupials reached Australia via Antarctica about 50 mya, shortly after Australia had split off suggesting a single dispersion event of just one species.[98] Evolutionary theory suggests that the Australian marsupials descended from the older ones found in the Americas. Geologic evidence suggests that between 30 and 40 million years ago South America and Australia were still part of the Southern Hemisphere super continent of Gondwana and that they were connected by land that is now part of Antarctica. Therefore, when combining the models, scientists could predict that marsupials migrated from what is now South America, through Antarctica, and then to present-day Australia between 40 and 30 million years ago. A first marsupial fossil of the extinct family Polydolopidae was found on Seymour Island on the Antarctic Peninsula in 1982.[99] Further fossils have subsequently been found, including members of the marsupial orders Didelphimorphia (opossum) and Microbiotheria,[100] as well as ungulates and a member of the enigmatic extinct order Gondwanatheria, possibly Sudamerica ameghinoi.[101][102][103]
Migration, isolation, and distribution of the camel
The history of the camel provides an example of how fossil evidence can be used to reconstruct migration and subsequent evolution. The fossil record indicates that the evolution of camelids started in North America (see figure 6c), from which, six million years ago, they migrated across the Bering Strait into Asia and then to Africa, and 3.5 million years ago through the Isthmus of Panama into South America. Once isolated, they evolved along their own lines, giving rise to the Bactrian camel and Dromedary in Asia and Africa and the llama and its relatives in South America. Camelids then went extinct in North America at the end of the last ice age.[104]
Evidence from selection
Examples for the evidence for evolution often stem from direct observation of natural selection in the field and the laboratory. Scientists have observed and documented a multitude of events where natural selection is in action. The most well known examples are antibiotic resistance in the medical field along with better-known laboratory experiments documenting evolution's occurrence. Natural selection is tantamount to common descent in that long-term occurrence and selection pressures can lead to the diversity of life on earth as found today. All adaptations—documented and undocumented changes concerned—are caused by natural selection (and a few other minor processes). The examples below are only a small fraction of the actual experiments and observations.
Specific examples of natural selection in the lab and in the field
Antibiotic and pesticide resistance
The development and spread of antibiotic-resistant bacteria, like the spread of pesticide-resistant forms of plants and insects, is evidence for evolution of species, and of change within species. Thus the appearance of vancomycin-resistant Staphylococcus aureus, and the danger it poses to hospital patients, is a direct result of evolution through natural selection. The rise of Shigella strains resistant to the synthetic antibiotic class of sulfonamides also demonstrates the generation of new information as an evolutionary process.[105] Similarly, the appearance of DDT resistance in various forms of Anopheles mosquitoes, and the appearance of myxomatosis resistance in breeding rabbit populations in Australia, are both evidence of the existence of evolution in situations of evolutionary selection pressure in species in which generations occur rapidly.
Artificial selection

Artificial selection demonstrates the diversity that can exist among organisms that share a relatively recent common ancestor. In artificial selection, one species is bred selectively at each generation, allowing only those organisms that exhibit desired characteristics to reproduce. These characteristics become increasingly well developed in successive generations. Artificial selection was successful long before science discovered the genetic basis. Examples of artificial selection would be dog breeding, genetically modified food, flower breeding, cultivation of foods such as wild cabbage,[106] and others.
Experimental evolution
Experimental evolution uses controlled experiments to test hypotheses and theories of evolution. In one early example, William Dallinger set up an experiment shortly before 1880, subjecting microbes to heat with the aim of forcing adaptive changes. His experiment ran for around seven years, and his published results were acclaimed, but he did not resume the experiment after the apparatus failed.[107]
A well-known and larger-scale example of experimental evolution is Richard Lenski's multi-generation experiment with Escherichia coli. Lenski observed that some strains of E. coli evolved a complex new ability, the ability to metabolize citrate, after tens of thousands of generations.[108][109] The evolutionary biologist Jerry Coyne commented as a critique of creationism, saying, "the thing I like most is it says you can get these complex traits evolving by a combination of unlikely events. That's just what creationists say can't happen."[108] In addition to the metabolic changes, the different bacterial populations were found to have diverged in respect to both morphology (the overall size of the cell) and fitness (of which was measured in competition with the ancestors).[110] The E. coli long-term evolution experiment that began in 1988 is still in progress, and has shown adaptations including the evolution of a strain of E. coli that was able to grow on citric acid in the growth media—a trait absent in all other known forms of E. coli, including the initial strain.
Guppies
While studying guppies (Poecilia reticulata) in Trinidad, biologist John Endler detected selection at work on the fish populations. To rule out alternative possibilities, Endler set up a highly controlled experiment to mimic the natural habitat by constructing ten ponds within a laboratory greenhouse at Princeton University. Each pond contained gravel to exactly match that of the natural ponds. After capturing a random sample of guppies from ponds in Trinidad, he raised and mixed them to create similar genetically diverse populations and measured each fish (spot length, spot height, spot area, relative spot length, relative spot height, total patch area, and standard body lengths). For the experiment he added Crenicichla alta (P. reticulata's main predator) in four of the ponds, Rivulus hartii (a non-predator fish) in four of the ponds, and left the remaining two ponds empty with only the guppies. After 10 generations, comparisons were made between each pond's guppy populations and measurements were taken again. Endler found that the populations had evolved dramatically different color patterns in the control and non-predator pools and drab color patterns in the predator pool. Predation pressure had caused a selection against standing out from background gravel.[111]
In parallel, during this experiment, Endler conducted a field experiment in Trinidad where he caught guppies from ponds where they had predators and relocated them to ponds upstream where the predators did not live. After 15 generations, Endler found that the relocated guppies had evolved dramatic and colorful patterns. Essentially, both experiments showed convergence due to similar selection pressures (i.e. predator selection against contrasting color patterns and sexual selection for contrasting color patterns).[111]
Additionally, in a later study by David Reznick, the field population was examined 11 years later after Endler relocated the guppies to high streams. The study found that the populations has evolved in a number of different ways: bright color patterns, late maturation, larger sizes, smaller litter sizes, and larger offspring within litters.[112]
Additional studies of P. reticulata and their predators in the streams of Trinidad have indicated that varying modes of selection through predation have not only changed the guppies color patterns, sizes, and behaviors, but their life histories and life history patterns.[113]
Humans
Natural selection is observed in contemporary human populations, with recent findings demonstrating that the population at risk of the severe debilitating disease kuru has significant over-representation of an immune variant of the prion protein gene G127V versus non-immune alleles. Scientists postulate one of the reasons for the rapid selection of this genetic variant is the lethality of the disease in non-immune persons.[114][115] Other reported evolutionary trends in other populations include a lengthening of the reproductive period, reduction in cholesterol levels, blood glucose and blood pressure.[116]
A well known example of selection occurring in human populations is lactose tolerance. Lactose intolerance is the inability to metabolize lactose, because of a lack of the required enzyme lactase in the digestive system. The normal mammalian condition is for the young of a species to experience reduced lactase production at the end of the weaning period (a species-specific length of time). In humans, in non-dairy consuming societies, lactase production usually drops about 90% during the first four years of life, although the exact drop over time varies widely.[117] Lactase activity persistence in adults is associated with two polymorphisms: C/T 13910 and G/A 22018 located in the MCM6 gene.[118] This gene difference eliminates the shutdown in lactase production, making it possible for members of these populations to continue consumption of raw milk and other fresh and fermented dairy products throughout their lives without difficulty. This appears to be an evolutionarily recent (around 10,000 years ago [and 7,500 years ago in Europe][119]) adaptation to dairy consumption,[120] and has occurred independently in both northern Europe and east Africa in populations with a historically pastoral lifestyle.[121][122]
Human-caused selection pressures in studied organisms
An example involving the direct observation of gene modification due to selection pressures is the resistance to PCBs in codfish. After General Electric dumped polychlorinated biphenyls (PCBs) in the Hudson River from 1947 through 1976, tomcods (Microgadus tomcod) living in the river were found to have evolved an increased resistance to the compound's toxic effects.[123]
The tolerance to the toxins is due to a change in the coding section of specific gene. Genetic samples were taken from the cods from 8 different rivers in the New England region: the St. Lawrence River, Miramichi River, Margaree River, Squamscott River, Niantic River, the Shinnecock Basic, the Hudson River, and the Hackensack River. Genetic analysis found that in the population of tomcods in the four southernmost rivers, the gene AHR2 (aryl hydrocarbon receptor 2) was present as an allele with a difference of two amino acid deletions.[124]
This deletion conferred a resistance to PCB in the fish species and was found in 99% of Hudson River tomcods, 92% in the Hackensack River, 6% in the Niantic River, and 5% in Shinnecock Bay.[124] This pattern along the sampled bodies of waters infers a direct correlation of selective pressures leading to the evolution of PCB resistance in Atlantic tomcod fish.[124]
A similar study was also done regarding the polycyclic aromatic hydrocarbons (PAHs) that pollute the waters of the Elizabeth River in Portsmouth, Virginia. This chemical is a product of creosote, a type of tar. The Atlantic killifish (Fundulus heteroclitus) has evolved a resistance to PAHs involving the AHR gene (the same gene involved in the tomcods). This particular study focused on the resistance to "acute toxicity and cardiac teratogenesis" caused by PAHs. that mutated within the tomcods in the Hudson River.[125]
Like the codfish, human-caused pollution can come in different forms. Radiotrophic fungi is a perfect example of natural selection taking place after a chemical accident. Radiotrophic fungi appears to use the pigment melanin to convert gamma radiation into chemical energy for growth[126][127] and were first discovered in 2007 as black molds growing inside and around the Chernobyl Nuclear Power Plant.[126] Research at the Albert Einstein College of Medicine showed that three melanin-containing fungi, Cladosporium sphaerospermum, Wangiella dermatitidis, and Cryptococcus neoformans, increased in biomass and accumulated acetate faster in an environment in which the radiation level was 500 times higher than in the normal environment.
Another example of organisms adapting to human-caused conditions are Nylon-eating bacteria: a strain of Flavobacterium that are capable of digesting certain byproducts of nylon 6 manufacturing. There is scientific consensus that the capacity to synthesize nylonase most probably developed as a single-step mutation that survived because it improved the fitness of the bacteria possessing the mutation. This is seen as a good example of evolution through mutation and natural selection that has been observed as it occurs and could not have come about until the production of nylon by humans.[128][129][130][131]
Urban wildlife is a broad and easily observable case of human-caused selection pressure on wildlife. With the growth in human habitats, different animals have adapted to survive within these urban environments. These types of environments can exert selection pressures on organisms, often leading to new adaptations. For example, the weed Crepis sancta, found in France, has two types of seed, heavy and fluffy. The heavy ones land nearby to the parent plant, whereas fluffy seeds float further away on the wind. In urban environments, seeds that float far often land on infertile concrete. Within about 5–12 generations, the weed evolves to produce significantly heavier seeds than its rural relatives.[132][133] Other examples of urban wildlife are rock pigeons and species of crows adapting to city environments around the world; African penguins in Simon's Town; baboons in South Africa; and a variety of insects living in human habitations. Studies have been conducted and have found striking changes to animals' (more specifically mammals') behavior and physical brain size due to their interactions with human-created environments.[134][135]
Other examples include phenotypic changes such as in the classic example of light-to-dark color adaptation in the peppered moth due to pollution from the Industrial Revolution in England.
Italian wall lizards
In 1971, ten adult specimens of Podarcis sicula (the Italian wall lizard) were transported from the Croatian island of Pod Kopište to the island Pod Mrčaru (about 3.5 km to the east). Both islands lie in the Adriatic Sea near Lastovo, where the lizards founded a new bottlenecked population.[136][137] The two islands have similar size, elevation, microclimate, and a general absence of terrestrial predators[137] and the P. sicula expanded for decades without human interference, even out-competing the (now locally extinct[136]) Podarcis melisellensis population.[138]
In the 1990s, scientists returned to Pod Mrčaru and found that the lizards currently occupying Mrčaru differ greatly from those on Kopište. While mitochondrial DNA analyses have verified that P. sicula currently on Mrčaru are genetically very similar to the Kopište source population,[136] the new Mrčaru population of P. sicula was described as having a larger average size, shorter hind limbs, lower maximal sprint speed and altered response to simulated predatory attacks compared to the original Kopište population.[137] These population changes in morphology and behavior were attributed to "relaxed predation intensity" and greater protection from vegetation on Mrčaru.[137]
In 2008, further analysis revealed that the Mrčaru population of P. sicula have significantly different head morphology (longer, wider, and taller heads) and increased bite force compared to the original Kopište population.[136] This change in head shape corresponded with a shift in diet: Kopište P. sicula are primarily insectivorous, but those on Mrčaru eat substantially more plant matter.[136] The changes in foraging style may have contributed to a greater population density and decreased territorial behavior of the Mrčaru population.[136]
Another difference found between the two populations was the discovery, in the Mrčaru lizards, of cecal valves, which slow down food passage and provide fermenting chambers, allowing commensal microorganisms to convert cellulose to nutrients digestible by the lizards.[136] Additionally, the researchers discovered that nematodes were common in the guts of Mrčaru lizards, but absent from Kopište P. sicula, which do not have cecal valves.[136] The cecal valves, which occur in less than 1 percent of all known species of scaled reptiles,[136] have been described as an "adaptive novelty, a brand new feature not present in the ancestral population and newly evolved in these lizards".[139]
Mimulus aurantiacus (monkeyflower) radiation
Both subspecies Mimulus aurantiacus puniceus (red-flowered) and Mimulus aurantiacus australis (yellow-flowered) of monkeyflowers are isolated due to the preferences of their hummingbird and hawkmoth pollinators. The radiation of M. aurantiacus subspecies are mostly yellow colored; however, both M. a. ssp. puniceus and M. a. ssp. flemingii are red. Phylogenetic analysis suggests two independent origins of red-colored flowers that arose due to cis-regulatory mutations in the gene MaMyb2 that is present in all M. aurantiacus subspecies. Further research suggested that two independent mutations did not take place, but one MaMyb2 allele was transferred via introgressive hybridization.[140] This study presents an example of the overlap of research in various disciplines. Gene isolation and cis-regulatory functions; phylogenetic analysis; geographic location and pollinator preference; and species hybridization and speciation are just some of the areas in which data can be obtained to document the occurrence of evolution.
Evidence from speciation
Speciation is the evolutionary process by which new biological species arise. Speciation can occur from a variety of different causes and are classified in various forms (e.g. allopatric, sympatric, polyploidization, etc.). Scientists have observed numerous examples of speciation in the laboratory and in nature, however, evolution has produced far more species than an observer would consider necessary. For example, there are well over 350,000 described species of beetles.[141] Great examples of observed speciation come from the observations of island biogeography and the process of adaptive radiation, both explained in an earlier section. Other examples of speciation can be found through paleontological studies of species within geologic strata. Paleontology cannot distinguish species on a genetic basis like research on extant species can; however, it can rely on various other methods to determine species differentiation. The examples described below represent different types of speciation observations and provide strong evidence for common descent. These examples are by no means exhaustive and are only a small fraction of the instances observed.
Specific examples
Blackcap
The bird species, Sylvia atricapilla, commonly referred to as blackcaps, lives in Germany and flies southwest to Spain while a smaller group flies northwest to Great Britain during the winter. Gregor Rolshausen from the University of Freiburg found that the genetic separation of the two populations is already in progress. The differences found have arisen in about 30 generations. With DNA sequencing, the individuals can be assigned to a correct group with an 85% accuracy. Stuart Bearhop from the University of Exeter reported that birds wintering in England tend to mate only among themselves, and not usually with those wintering in the Mediterranean.[142] It is still inference to say that the populations will become two different species, but researchers expect it due to the continued genetic and geographic separation.[143]
Drosophila melanogaster
.jpg)
William R. Rice and George W. Salt found experimental evidence of sympatric speciation in the common fruit fly. They collected a population of Drosophila melanogaster from Davis, California and placed the pupae into a habitat maze. Newborn flies had to investigate the maze to find food. The flies had three choices to take in finding food. Light and dark (phototaxis), up and down (geotaxis), and the scent of acetaldehyde and the scent of ethanol (chemotaxis) were the three options. This eventually divided the flies into 42 spatio-temporal habitats.
They then cultured two strains that chose opposite habitats. One of the strains emerged early, immediately flying upward in the dark attracted to the acetaldehyde. The other strain emerged late and immediately flew downward, attracted to light and ethanol. Pupae from the two strains were then placed together in the maze and allowed to mate at the food site. They then were collected. A selective penalty was imposed on the female flies that switched habitats. This entailed that none of their gametes would pass on to the next generation. After 25 generations of this mating test, it showed reproductive isolation between the two strains. They repeated the experiment again without creating the penalty against habitat switching and the result was the same; reproductive isolation was produced.[144][145][146]
Gall wasps
A study of the gall-forming wasp species Belonocnema treatae found that populations inhabiting different host plants (Quercus geminata and Q. Virginiana) exhibited different body size and gall morphology alongside an strong expression of sexual isolation. The study hypothesized that B. treatae populations inhabiting different host plants would show evidence of divergent selection promoting speciation. The researchers sampled gall wasp species and oak tree localities, measured body size (right hand tibia of each wasp), and counted gall chamber numbers. In addition to measurements, they conducted mating assays and statistical analyses. Genetic analysis was also conducted on two mtDNA sites (416 base pairs from cytochrome C and 593 base pairs from cytochrome oxidase ) to “control for the confounding effects of time since divergence among allopatric populations”.[147]
In an additional study, the researchers studied two gall wasp species B. treatae and Disholcaspis quercusvirens and found strong morphological and behavioral variation among host-associated populations. This study further confounded prerequisites to speciation.[148]
Hawthorn fly
One example of evolution at work is the case of the hawthorn fly, Rhagoletis pomonella, also known as the apple maggot fly, which appears to be undergoing sympatric speciation.[149] Different populations of hawthorn fly feed on different fruits. A distinct population emerged in North America in the 19th century some time after apples, a non-native species, were introduced. This apple-feeding population normally feeds only on apples and not on the historically preferred fruit of hawthorns. The current hawthorn feeding population does not normally feed on apples. Some evidence, such as the fact that six out of thirteen allozyme loci are different, that hawthorn flies mature later in the season and take longer to mature than apple flies; and that there is little evidence of interbreeding (researchers have documented a 4–6% hybridization rate) suggests that speciation is occurring.[150][151][152][153][154]
London Underground mosquito
The London Underground mosquito is a species of mosquito in the genus Culex found in the London Underground. It evolved from the overground species Culex pipiens.
This mosquito, although first discovered in the London Underground system, has been found in underground systems around the world. It is suggested that it may have adapted to human-made underground systems since the last century from local above-ground Culex pipiens,[155] although more recent evidence suggests that it is a southern mosquito variety related to Culex pipiens that has adapted to the warm underground spaces of northern cities.[156]
The species have very different behaviours,[157] are extremely difficult to mate,[155] and with different allele frequency, consistent with genetic drift during a founder event.[158] More specifically, this mosquito, Culex pipiens molestus, breeds all-year round, is cold intolerant, and bites rats, mice, and humans, in contrast to the above ground species Culex pipiens that is cold tolerant, hibernates in the winter, and bites only birds. When the two varieties were cross-bred the eggs were infertile suggesting reproductive isolation.[155][157]
The genetic data indicates that the molestus form in the London Underground mosquito appears to have a common ancestry, rather than the population at each station being related to the nearest aboveground population (i.e. the pipiens form). Byrne and Nichols' working hypothesis was that adaptation to the underground environment had occurred locally in London only once.
These widely separated populations are distinguished by very minor genetic differences, which suggest that the molestus form developed: a single mtDNA difference shared among the underground populations of ten Russian cities;[159] a single fixed microsatellite difference in populations spanning Europe, Japan, Australia, the middle East and Atlantic islands.[156]
Mollies
The shortfin molly (Poecilia mexicana) is a small fish that lives in the Sulfur Caves of Mexico. Years of study on the species have found that two distinct populations of mollies—the dark interior fish and the bright surface water fish—are becoming more genetically divergent.[160] The populations have no obvious barrier separating the two; however, it was found that the mollies are hunted by a large water bug (Belostoma spp). Tobler collected the bug and both types of mollies, placed them in large plastic bottles, and put them back in the cave. After a day, it was found that, in the light, the cave-adapted fish endured the most damage, with four out of every five stab-wounds from the water bugs sharp mouthparts. In the dark, the situation was the opposite. The mollies' senses can detect a predator's threat in their own habitats, but not in the other ones. Moving from one habitat to the other significantly increases the risk of dying. Tobler plans on further experiments, but believes that it is a good example of the rise of a new species.[161]
Polar bear
A remarkable example of natural selection, geographic isolation, and speciation in progress is the relationship between the polar bear (Ursus maritimus) and the brown bear (Ursus arctos). Considered separate species throughout their ranges;[162] however, it has been documented that they possess the capability to interbreed and produce fertile offspring. This introgressive hybridization has occurred both in the wild and in captivity and has been documented[163] and verified with DNA testing.[164] The oldest known fossil evidence of polar bears dates around 130,000 to 110,000 years ago;[165] however, molecular data has revealed varying estimates of divergence time. Mitochondrial DNA analysis has given an estimate of 150,000 years ago[165] while nuclear genome analysis has shown an approximate divergence of 603,000 years ago.[166] Recent research using the complete genomes (rather than mtDNA or partial nuclear genomes) establishes the divergence of polar and brown bears between 479-343 thousand years ago.[167] Despite the differences in divergence rates, molecular research suggests the sister species have undergone a highly complex process of speciation and admixture between the two.[168]
Polar bears have acquired significant anatomical and physiological differences from the brown bear that allow it to comfortably survive in conditions that the brown bear likely could not. Notable examples include the ability to swim sixty miles or more at a time in freezing waters, fur that blends with the snow, and to stay warm in the arctic environment, an elongated neck that makes it easier to keep their heads above water while swimming, and oversized and heavy-matted webbed feet that act as paddles when swimming. It has also evolved small papillae and vacuole-like suction cups on the soles to make them less likely to slip on the ice, alongside smaller ears for a reduction of heat loss, eyelids that act like sunglasses, accommodations for their all-meat diet, a large stomach capacity to enable opportunistic feeding, and the ability to fast for up to nine months while recycling their urea.[169][170] This example presents a macro-evolutionary change involving an amalgamation of several fields of evolutionary biology, e.g. adaptation through natural selection, geographic isolation, speciation, and hybridization.
Thale cress

Kirsten Bomblies et al. from the Max Planck Institute for Developmental Biology discovered that two genes passed down by each parent of the thale cress plant, Arabidopsis thaliana. When the genes are passed down, it ignites a reaction in the hybrid plant that turns its own immune system against it. In the parents, the genes were not detrimental, but they evolved separately to react defectively when combined.[171]
To test this, Bomblies crossed 280 genetically different strains of Arabidopsis in 861 distinct ways and found that 2 per cent of the resulting hybrids were necrotic. Along with allocating the same indicators, the 20 plants also shared a comparable collection of genetic activity in a group of 1,080 genes. In almost all of the cases, Bomblies discovered that only two genes were required to cause the autoimmune response. Bomblies looked at one hybrid in detail and found that one of the two genes belonged to the NB-LRR class, a common group of disease resistance genes involved in recognizing new infections. When Bomblies removed the problematic gene, the hybrids developed normally.[171]
Over successive generations, these incompatibilities could create divisions between different plant strains, reducing their chances of successful mating and turning distinct strains into separate species.[172]
Turborotalia
Seafloor sediments can provide a significant amount of data concerning planktonic microfossils. The succession of fossils in stratigraphy can be used to determine evolutionary trends among fossil organisms. In addition, incidences of speciation can be interpreted from the data and numerous studies have been conducted documenting both morphological evolution and speciation. A recent study was conducted involving the planktonic foraminifer Turborotalia—one of the largest of its kind. The authors Paul N. Pearson and Thomas H. G. Ezard extracted “51 stratigraphically ordered samples from a site within the oceanographically stable tropical North Pacific gyre”. Two hundred individual species were examined on each of the 51 samples using ten specific morphological traits (size, compression index, chamber aspect ratio, chamber inflation, aperture aspect ratio, test height, test expansion, umbilical angle, coiling direction, and the number of chambers in the final whorl).
Utilizing multivariate statistical clustering methods, the study found that the species continued to evolve non-directionally within the Eocene from 45 Ma to about 36 Ma. However, from 36 Ma to approximately 34 Ma, the stratigraphic layers showed two distinct clusters with significantly defining characteristics distinguishing one another from a single species. The authors concluded that speciation must have occurred and that the two new species were ancestral to the prior species.[173] Just as in most of evolutionary biology, this example represents the interdisciplinary nature of the field and the necessary collection of data from various fields (e.g. oceanography, paleontology) and the integration of mathematical analysis (e.g. biometry).
Interspecies fertility or hybridization
Understood from laboratory studies and observed instances of speciation in nature, finding species that are able reproduce successfully or create hybrids between two different species infers that their relationship is close. In conjunction with this, hybridization has been found to be a precursor to the creation of new species by being a source of new genes for a species. The examples provided are only a small fraction of the observed instances of speciation through hybridization. Plants are often subject to the creation of a new species though hybridization.
Mimulus peregrinus
The creation of a new allopolyploid species (Mimulus peregrinus) was observed on the banks of the Shortcleuch Water—a river in Leadhills, South Lanarkshire, Scotland. Parented from the cross of the two species Mimulus guttatus (containing 14 pairs of chromosomes) and Mimulus luteus (containing 30-31 pairs from a chromosome duplication), M. peregrinus has six copies of its chromosomes (caused by the duplication of the sterile hybrid triploid). Due to the nature of these species, they have the ability to self-fertilize. Because of its number of chromosomes it is not able to pair with M. guttatus, M. luteus, or their sterile triploid offspring. M. peregrinus will either die producing no offspring or reproduce with itself making a new species.[174][175]
Raphanobrassica
Raphanobrassica includes all intergeneric hybrids between the genera Raphanus (radish) and Brassica (cabbages, etc.).[176][177]
The Raphanobrassica is an allopolyploid cross between the radish (Raphanus sativus) and cabbage (Brassica oleracea). Plants of this parentage are now known as radicole. Two other fertile forms of Raphanobrassica are known. Raparadish, an allopolyploid hybrid between Raphanus sativus and Brassica rapa is grown as a fodder crop. "Raphanofortii" is the allopolyploid hybrid between Brassica tournefortii and Raphanus caudatus.
The Raphanobrassica is a fascinating plant, because (in spite of its hybrid nature), it is not sterile. This has led some botanists to propose that the accidental hybridization of a flower by pollen of another species in nature could be a mechanism of speciation common in higher plants.
Senecio (groundsel)
The Welsh groundsel is an allopolyploid, a plant that contains sets of chromosomes originating from two different species. Its ancestor was Senecio × baxteri, an infertile hybrid that can arise spontaneously when the closely related groundsel (Senecio vulgaris) and Oxford ragwort (Senecio squalidus) grow alongside each other. Sometime in the early 20th century, an accidental doubling of the number of chromosomes in an S. × baxteri plant led to the formation of a new fertile species.[178][179]
The York groundsel (Senecio eboracensis) is a hybrid species of the self-incompatible Senecio squalidus (also known as Oxford ragwort) and the self-compatible Senecio vulgaris (also known as common groundsel). Like S. vulgaris, S. eboracensis is self-compatible, however, it shows little or no natural crossing with its parent species, and is therefore reproductively isolated, indicating that strong breed barriers exist between this new hybrid and its parents.
It resulted from a backcrossing of the F1 hybrid of its parents to S. vulgaris. S. vulgaris is native to Britain, while S. squalidus was introduced from Sicily in the early 18th century; therefore, S. eboracensis has speciated from those two species within the last 300 years.
Other hybrids descended from the same two parents are known. Some are infertile, such as S. x baxteri. Other fertile hybrids are also known, including S. vulgaris var. hibernicus, now common in Britain, and the allohexaploid S. cambrensis, which according to molecular evidence probably originated independently at least three times in different locations. Morphological and genetic evidence support the status of S. eboracensis as separate from other known hybrids.[180]
Tragopogon (salsify)

Tragopogon is one example where hybrid speciation has been observed. In the early 20th century, humans introduced three species of salsify into North America. These species, the western salsify (Tragopogon dubius), the meadow salsify (Tragopogon pratensis), and the oyster plant (Tragopogon porrifolius), are now common weeds in urban wastelands. In the 1950s, botanists found two new species in the regions of Idaho and Washington, where the three already known species overlapped. One new species, Tragopogon miscellus, is a tetraploid hybrid of T. dubius and T. pratensis. The other new species, Tragopogon mirus, is also an allopolyploid, but its ancestors were T. dubius and T. porrifolius. These new species are usually referred to as "the Ownbey hybrids" after the botanist who first described them. The T. mirus population grows mainly by reproduction of its own members, but additional episodes of hybridization continue to add to the T. mirus population.[181]
T. dubius and T. pratensis mated in Europe but were never able to hybridize. A study published in March 2011 found that when these two plants were introduced to North America in the 1920s, they mated and doubled the number of chromosomes in there hybrid Tragopogon miscellus allowing for a "reset" of its genes, which in turn, allows for greater genetic variation. Professor Doug Soltis of the University of Florida said, "We caught evolution in the act…New and diverse patterns of gene expression may allow the new species to rapidly adapt in new environments".[182][183] This observable event of speciation through hybridization further advances the evidence for the common descent of organisms and the time frame in which the new species arose in its new environment. The hybridizations have been reproduced artificially in laboratories from 2004 to present day.
Evidence from computation and mathematical iteration
Computer science allows the iteration of self-changing complex systems to be studied, allowing a mathematical understanding of the nature of the processes behind evolution; providing evidence for the hidden causes of known evolutionary events. The evolution of specific cellular mechanisms like spliceosomes that can turn the cell's genome into a vast workshop of billions of interchangeable parts that can create tools that create tools that create tools that create us can be studied for the first time in an exact way.
"It has taken more than five decades, but the electronic computer is now powerful enough to simulate evolution,"[184] assisting bioinformatics in its attempt to solve biological problems.
Computational evolutionary biology has enabled researchers to trace the evolution of a large number of organisms by measuring changes in their DNA, rather than through physical taxonomy or physiological observations alone. It has compared entire genomes permitting the study of more complex evolutionary events, such as gene duplication, horizontal gene transfer, and the prediction of factors important in speciation. It has also helped build complex computational models of populations to predict the outcome of the system over time and track and share information on an increasingly large number of species and organisms.
Future endeavors are to reconstruct a now more complex tree of life.
Christoph Adami, a professor at the Keck Graduate Institute made this point in Evolution of biological complexity:
- To make a case for or against a trend in the evolution of complexity in biological evolution, complexity must be both rigorously defined and measurable. A recent information-theoretic (but intuitively evident) definition identifies genomic complexity with the amount of information a sequence stores about its environment. We investigate the evolution of genomic complexity in populations of digital organisms and monitor in detail the evolutionary transitions that increase complexity. We show that, because natural selection forces genomes to behave as a natural "Maxwell Demon", within a fixed environment, genomic complexity is forced to increase.[185]
David J. Earl and Michael W. Deem—professors at Rice University made this point in Evolvability is a selectable trait:
- Not only has life evolved, but life has evolved to evolve. That is, correlations within protein structure have evolved, and mechanisms to manipulate these correlations have evolved in tandem. The rates at which the various events within the hierarchy of evolutionary moves occur are not random or arbitrary but are selected by Darwinian evolution. Sensibly, rapid or extreme environmental change leads to selection for greater evolvability. This selection is not forbidden by causality and is strongest on the largest-scale moves within the mutational hierarchy. Many observations within evolutionary biology, heretofore considered evolutionary happenstance or accidents, are explained by selection for evolvability. For example, the vertebrate immune system shows that the variable environment of antigens has provided selective pressure for the use of adaptable codons and low-fidelity polymerases during somatic hypermutation. A similar driving force for biased codon usage as a result of productively high mutation rates is observed in the hemagglutinin protein of influenza A.[186]
"Computer simulations of the evolution of linear sequences have demonstrated the importance of recombination of blocks of sequence rather than point mutagenesis alone. Repeated cycles of point mutagenesis, recombination, and selection should allow in vitro molecular evolution of complex sequences, such as proteins."[187] Evolutionary molecular engineering, also called directed evolution or in vitro molecular evolution involves the iterated cycle of mutation, multiplication with recombination, and selection of the fittest of individual molecules (proteins, DNA, and RNA). Natural evolution can be relived showing us possible paths from catalytic cycles based on proteins to based on RNA to based on DNA.[187][188][189][190]
Specific examples
Avida simulation
Richard Lenski, Charles Ofria, et al. at Michigan State University developed an artificial life computer program with the ability to detail the evolution of complex systems. The system uses values set to determine random mutations and allows for the effect of natural selection to conserve beneficial traits. The program was dubbed Avida and starts with an artificial petri dish where organisms reproduce and perform mathematical calculations to acquire rewards of more computer time for replication. The program randomly adds mutations to copies of the artificial organisms to allow for natural selection. As the artificial life reproduced, different lines adapted and evolved depending on their set environments. The beneficial side to the program is that it parallels that of real life at rapid speeds.[191][192][193]
See also
References
- ↑ Mount, D.M. (2004). Bioinformatics: Sequence and Genome Analysis (2nd ed.). Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY. ISBN 0-87969-608-7.
- ↑ Futuyma, Douglas J. (1998). Evolutionary Biology (3rd ed.). Sinauer Associates Inc. pp. 108–110. ISBN 0-87893-189-9.
- ↑ Haszprunar (1995). "The mollusca: Coelomate turbellarians or mesenchymate annelids?". In Taylor. Origin and evolutionary radiation of the Mollusca : centenary symposium of the Malacological Society of London. Oxford: Oxford Univ. Press. ISBN 0-19-854980-6.
- ↑ Kozmik, Z; Daube, M; Frei, E; Norman, B; Kos, L; Dishaw, LJ; Noll, M; Piatigorsky, J (2003). "Role of Pax genes in eye evolution: A cnidarian PaxB gene uniting Pax2 and Pax6 functions" (PDF). Developmental Cell 5 (5): 773–85. doi:10.1016/S1534-5807(03)00325-3. PMID 14602077.
- ↑ Land, M.F. and Nilsson, D.-E., Animal Eyes, Oxford University Press, Oxford (2002) ISBN 0-19-850968-5.
- ↑ Chen, F.C.; Li, W.H. (2001). "Genomic Divergences between Humans and Other Hominoids and the Effective Population Size of the Common Ancestor of Humans and Chimpanzees". American Journal of Human Genetics 68 (2): 444–56. doi:10.1086/318206. PMC 1235277. PMID 11170892.
- ↑ Cooper, G.M.; Brudno, M.; Green, E.D.; Batzoglou, S.; Sidow, A. (2003). "Quantitative Estimates of Sequence Divergence for Comparative Analyses of Mammalian Genomes". Genome Res. 13 (5): 813–20. doi:10.1101/gr.1064503. PMC 430923. PMID 12727901.
- ↑ The picture labeled "Human Chromosome 2 and its analogs in the apes" in the article Comparison of the Human and Great Ape Chromosomes as Evidence for Common Ancestry is literally a picture of a link in humans that links two separate chromosomes in the nonhuman apes creating a single chromosome in humans. Also, while the term originally referred to fossil evidence, this too is a trace from the past corresponding to some living beings that, when alive, physically embodied this link.
- ↑ The New York Times report Still Evolving, Human Genes Tell New Story, based on A Map of Recent Positive Selection in the Human Genome, states the International HapMap Project is "providing the strongest evidence yet that humans are still evolving" and details some of that evidence.
- ↑ Alberts, Bruce; Johnson, Alexander; Lewis, Julian; Raff, Martin; Roberts, Keith; Walter, Peter (March 2002). Molecular Biology of the Cell (4th ed.). Routledge. ISBN 0-8153-3218-1.
- ↑ "29+ Evidences for Macroevolution: The Scientific Case for Common Descent". Theobald, Douglas. Retrieved 2011-03-10.
- ↑ Van Der Kuyl, AC; Dekker, JT; Goudsmit, J (1999). "Discovery of a New Endogenous Type C Retrovirus (FcEV) in Cats: Evidence for RD-114 Being an FcEVGag-Pol/Baboon Endogenous Virus BaEVEnv Recombinant". Journal of Virology 73 (10): 7994–8002. PMC 112814. PMID 10482547.
- ↑ Sverdlov ED (February 2000). "Retroviruses and primate evolution". BioEssays 22 (2): 161–71. doi:10.1002/(SICI)1521-1878(200002)22:2<161::AID-BIES7>3.0.CO;2-X. PMID 10655035.
- ↑ Belshaw R, Pereira V, Katzourakis A; et al. (April 2004). "Long-term reinfection of the human genome by endogenous retroviruses". Proceedings of the National Academy of Sciences of the United States of America 101 (14): 4894–9. Bibcode:2004PNAS..101.4894B. doi:10.1073/pnas.0307800101. PMC 387345. PMID 15044706.
- ↑ Bonner TI, O'Connell C, Cohen M (August 1982). "Cloned endogenous retroviral sequences from human DNA". Proceedings of the National Academy of Sciences of the United States of America 79 (15): 4709–13. Bibcode:1982PNAS...79.4709B. doi:10.1073/pnas.79.15.4709. PMC 346746. PMID 6181510.
- ↑ "Converging Evidence for Evolution." Phylointelligence: Evolution for Everyone. Web. 26 Nov. 2010.
- ↑ Petrov DA, Hartl DL (2000). "Pseudogene evolution and natural selection for a compact genome". The Journal of Heredity 91 (3): 221–7. doi:10.1093/jhered/91.3.221. PMID 10833048.
- ↑ Okamoto, N.; Inouye, I. (2005). "A secondary symbiosis in progress". Science 310 (5746): 287. doi:10.1126/science.1116125. PMID 16224014.
- ↑ Okamoto, N.; Inouye, I. (2006). "Hatena arenicola gen. et sp. nov., a katablepharid undergoing probable plastid acquisition". Protist 157 (4): 401–19. doi:10.1016/j.protis.2006.05.011. PMID 16891155.
- ↑ Human Chromosome 2 is a fusion of two ancestral chromosomes by Alec MacAndrew; accessed 18 May 2006.
- ↑ Evidence of Common Ancestry: Human Chromosome 2 (video) 2007
- ↑ Yunis, J.J.; Prakash, O. (1982). "The origin of man: a chromosomal pictorial legacy". Science 215 (4539): 1525–1530. Bibcode:1982Sci...215.1525Y. doi:10.1126/science.7063861. PMID 7063861.
- ↑ Human and Ape Chromosomes; accessed 8 September 2007.
- ↑ Avarello, Rosamaria; Pedicini, A; Caiulo, A; Zuffardi, O; Fraccaro, M (1992). "Evidence for an ancestral alphoid domain on the long arm of human chromosome 2". Human Genetics 89 (2): 247–9. doi:10.1007/BF00217134. PMID 1587535.
- 1 2 Ijdo, J. W.; Baldini, A; Ward, DC; Reeders, ST; Wells, RA (1991). "Origin of human chromosome 2: an ancestral telomere-telomere fusion". Proceedings of the National Academy of Sciences 88 (20): 9051–5. Bibcode:1991PNAS...88.9051I. doi:10.1073/pnas.88.20.9051. PMC 52649. PMID 1924367.
- 1 2 Amino acid sequences in cytochrome c proteins from different species, adapted from Strahler, Arthur; Science and Earth History, 1997. page 348.
- ↑ Lurquin, P.F.; Stone, L. (2006). Genes, Culture, and Human Evolution: A Synthesis. Blackwell Publishing, Incorporated. p. 79. ISBN 1-4051-5089-0.
- ↑ 29+ Evidences for Macroevolution; Protein functional redundancy, Douglas Theobald, Ph.D.
- ↑ Castresana, J. (2001). "Cytochrome b Phylogeny and the Taxonomy of Great Apes and Mammals". Molecular Biology and Evolution 18 (4): 465–471. doi:10.1093/oxfordjournals.molbev.a003825. PMID 11264397.
- ↑ Pallen, Mark (2009). Rough Guide to Evolution. Rough Guides. pp. 200–206. ISBN 978-1-85828-946-5.
- ↑ Tanaka G, Hou X, Ma X, Edgecombe GD, Strausfeld NJ (October 2013). "Chelicerate neural ground pattern in a Cambrian great appendage arthropod". Nature 502 (7471): 364–7. Bibcode:2013Natur.502..364T. doi:10.1038/nature12520. PMID 24132294.
- 1 2 "29+ Evidences for Macroevolution: Part 2". TalkOrigins Archive. Retrieved 2006-11-08.
- ↑ Lambert, Katie. (2007-10-29) HowStuffWorks "How Atavisms Work". Animals.howstuffworks.com. Retrieved on 2011-12-06.
- ↑ JPG image
- ↑ Evolutionary Atavisms. Edwardtbabinski.us. Retrieved on 2011-12-06.
- ↑ Andrews, Roy Chapman (June 3, 1921). "A REMARKABLE CASE OF EXTERNAL HIND LIMBS IN A HUMPBACK WHALE" (PDF). American Museum Novitates.
- ↑ Tyson R, Graham JP, Colahan PT, Berry CR (2004). "Skeletal atavism in a miniature horse". Veterinary Radiology & Ultrasound : the Official Journal of the American College of Veterinary Radiology and the International Veterinary Radiology Association 45 (4): 315–7. doi:10.1111/j.1740-8261.2004.04060.x. PMID 15373256.
- ↑ Biello, David (2006-02-22). "Mutant Chicken Grows Alligatorlike Teeth". Scientific American. Retrieved 2009-03-08.
- ↑ Domes K, Norton RA, Maraun M, Scheu S (April 2007). "Reevolution of sexuality breaks Dollo's law". Proceedings of the National Academy of Sciences of the United States of America 104 (17): 7139–44. Bibcode:2007PNAS..104.7139D. doi:10.1073/pnas.0700034104. PMC 1855408. PMID 17438282.
- ↑ Held, Lewis I. (2010). "The Evo-Devo Puzzle of Human Hair Patterning". Evolutionary Biology 37 (2–3): 113–122. doi:10.1007/s11692-010-9085-4.
- 1 2 Futuyma, Douglas J. (1998). Evolutionary Biology (3rd ed.). Sinauer Associates Inc. p. 122. ISBN 0-87893-189-9.
- 1 2 29+ Evidences for Macroevolution: Part 1. Talkorigins.org. Retrieved on 2011-12-06.
- ↑ Coyne, Jerry A. (2009). Why Evolution is True. Viking. pp. 8–11. ISBN 978-0-670-02053-9.
- ↑ Darwin, Charles (1859). On the Origin of Species. John Murray. p. 420.
- ↑ Tuomi, J. (1981). "Structure and dynamics of Darwinian evolutionary theory" (PDF). Syst. Zool. 30 (1): 22–31. doi:10.2307/2992299.
- ↑ Aravind L, Iyer LM, Anantharaman V (2003). "The two faces of Alba: the evolutionary connection between proteins participating in chromatin structure and RNA metabolism". Genome Biology 4 (10): R64. doi:10.1186/gb-2003-4-10-r64. PMC 328453. PMID 14519199.
- ↑ Brochu, C. A.; Wagner, J. R.; Jouve, S.; Sumrall, C. D.; Densmore, L. D. (2009). "A correction corrected:Consensus over the meaning of Crocodylia and why it matters" (PDF). Syst. Biol. 58 (5): 537–543. doi:10.1093/sysbio/syp053. PMID 20525607.
- ↑ Bock, W. J. (2007). "Explanations in evolutionary theory" (PDF). J Zool Syst Evol Res 45 (2): 89–103. doi:10.1111/j.1439-0469.2007.00412.x.
- ↑ Kluge 2000
- ↑ Laurin 2000
- ↑ Fitzhugh 2006, p. 31
- ↑ Kluge 2000, p. 432
- ↑ Slifkin, Natan (2006). The Challenge of Creation... Zoo Torah. pp. 258–9. ISBN 1-933143-15-0.
- ↑ Coyne, Jerry A. (2009). Why Evolution Is True. Viking. pp. 69–70. ISBN 978-0-670-02053-9.
- ↑ West-Eberhard, Mary Jane (2003). Developmental plasticity and evolution. Oxford University Press. p. 232. ISBN 0-19-512234-8.
- ↑ "Example 1: Living whales and dolphins found with hindlimbs". Douglas Theobald. Retrieved 2011-03-20.
- ↑ Ridley, Mark (2004). Evolution (3rd ed.). Blackwell Publishing. p. 282. ISBN 1-4051-0345-0.
- 1 2 Dawkins, Richard (2009). The Greatest Show on Earth: The Evidence for Evolution. Bantam Press. pp. 364–365. ISBN 978-1-4165-9478-9.
- ↑ Williams, G.C. (1992). Natural selection: domains, levels, and challenges. Oxford Press. ISBN 0-19-506932-3.
- ↑ Coyne, Jerry A. (2009). Why Evolution is True. Viking. pp. 26–28. ISBN 978-0-670-02053-9.
- ↑ "Confessions of a Darwinist". Niles Eldredge. Retrieved 2010-06-22.
- ↑ Laboratory 11 – Fossil Preservation, by Pamela J. W. Gore, Georgia Perimeter College
- ↑ "Frequently Asked Questions". The Natural History Museum of Los Angeles County Foundation. Retrieved 2011-02-21.
- ↑ Dean, William Richard John; Milton, Suzanne Jane (1999). The Karoo: Ecological Patterns and Processes. Cambridge University Press. p. 31. ISBN 0-521-55450-0.
- ↑ Schadewald, Robert J. (1982). "Six "Flood" Arguments Creationists Can't Answer". Creation Evolution Journal 3: 12–17.
- ↑ "Obviously vertebrates must have had ancestors living in the Cambrian, but they were assumed to be invertebrate forerunners of the true vertebrates — protochordates. Pikaia has been heavily promoted as the oldest fossil protochordate." Richard Dawkins 2004 The Ancestor's Tale Page 289, ISBN 0-618-00583-8
- ↑ Chen, J. Y.; Huang, D. Y.; Li, C. W. (1999). "An early Cambrian craniate-like chordate". Nature 402 (6761): 518–522. Bibcode:1999Natur.402..518C. doi:10.1038/990080.
- ↑ Shu, D. G.; Morris, S. C.; Han, J.; Zhang, Z. F.; Yasui, K.; Janvier, P.; Chen, L.; Zhang, X. L.; Liu, J. N.; Li, Y.; Liu, H. -Q. (2003), "Head and backbone of the Early Cambrian vertebrate Haikouichthys", Nature 421 (6922): 526–529, Bibcode:2003Natur.421..526S, doi:10.1038/nature01264, PMID 12556891
- ↑ Legendre, Serge (1989). Les communautés de mammifères du Paléogène (Eocène supérieur et Oligocène) d'Europe occidentale : structures, milieux et évolution. München: F. Pfeil. p. 110. ISBN 978-3-923871-35-3.
- ↑ Academy of Natural Sciences - Joseph Leidy - American Horses
- ↑ "Shubin, Neil (2008). Your Inner Fish. Pantheon. ISBN 978-0-375-42447-2.
- ↑ Niedzwiedzki, G.; Szrek, P.; Narkiewicz, K.; Narkiewicz, M.; Ahlberg, P. (2010). "Tetrapod trackways from the early Middle Devonian period of Poland". Nature 463 (7227): 43–48. Bibcode:2010Natur.463...43N. doi:10.1038/nature08623. PMID 20054388.
- ↑ Cota-Sánchez, J. Hugo; Bomfim-Patrício, Márcia C. (2010). "Seed morphology, polyploidy and the evolutionary history of the epiphytic cactus Rhipsalis baccifera (Cactaceae)" (PDF). Polibotanica 29: 107–129. Retrieved 2013-02-28.
- ↑ Menkhorst, Peter; Knight, Frank (2001). A Field Guide to the Mammals of Australia. Oxford University Press. p. 14. ISBN 0-19-550870-X.
- ↑ Augee, Michael; Gooden, Brett; Musser, Anne (2006). Echidna: Extraordinary egg-laying mammal. CSIRO Publishing.
- ↑ "Polar Bears/Habitat & Distribution". SeaWorld Parks & Entertainment. Retrieved 2011-02-21.
- ↑ "Sirenians of the World". Save the Manatee Club. Retrieved 2011-02-21.
- ↑ Continental Drift and Evolution. Biology.clc.uc.edu (2001-03-25). Retrieved on 2011-12-06.
- 1 2 3 4 Coyne, Jerry A. (2009). Why Evolution is True. Viking. pp. 99–110. ISBN 978-0-670-02053-9.
- ↑ Murphy, James B.; Ciofi, Claudio; de la Panouse, Colomba; Walsh, Trooper, eds. (2002). Komodo Dragons: Biology and Conservation (Zoo and Aquarium Biology and Conservation Series). Washington, D.C.: Smithsonian Books. ISBN 1-58834-073-2.
- ↑ Burdick, Alan (2007-03-25). "The Wonder Land of Socotra, Yemen". ALAN BURDICK. Retrieved 2010-07-08.
- ↑ "Tuatara". New Zealand Ecology: Living Fossils. TerraNature Trust. 2004. Retrieved 2006-11-10.
- ↑ "Facts about tuatara". Conservation: Native Species. Threatened Species Unit, Department of Conservation, Government of New Zealand. Retrieved 2007-02-10.
- ↑ "New Caledonia's most wanted". Retrieved 2010-07-08.
- ↑ "Giant bushy-tailed cloud rat (Crateromys schadenbergi)". Retrieved 2010-07-08.
- ↑ Rabor, D.S. (1986). Guide to Philippine Flora and Fauna. Natural Resources Management Centre, Ministry of Natural Resources and University of the Philippines.
- ↑ Robert R. Humphrey. The Boojum and its Home
- ↑ Schofield, James (27 July 2001). "Lake Baikal's Vanishing Nerpa Seal". The Moscow Times. Retrieved 2007-09-27.
- ↑ Baldwin, B. G.; Robichaux, R. H. (1995). "Historical biogeography and ecology of the Hawaiian silversword alliance (Asteraceae). New molecular phylogenetic perspectives". In Wagner, W. L.; Funk, V. A. Hawaiian biogeography: evolution on a hotspot archipelago. Washington: Smithsonian Institution Press. pp. 259–287.
- ↑ "Adaptive Radiation and Hybridization in the Hawaiian Silversword Alliance". University of Hawaii Botany Department.
- ↑ Pallen, Mark (2009). Rough Guide to Evolution. Rough Guides. p. 87. ISBN 978-1-85828-946-5.
- ↑ Evolution – A-Z – Ring species. Blackwellpublishing.com. Retrieved on 2011-12-06.
- ↑ Discovering a ring species. Evolution.berkeley.edu. Retrieved on 2011-12-06.
- ↑ Davis, Paul and Kenrick, Paul. 2004. Fossil Plants. Smithsonian Books (in association with the Natural History Museum of London), Washington, D.C. ISBN 1-58834-156-9
- ↑ Coyne, Jerry A. (2009). Why Evolution is True. Viking. p. 103. ISBN 978-0-670-02053-9.
- ↑ Pioneer Productions (2010-01-19). "Episode Guide". How The Earth Was Made. Season 2. Episode 8. History channel.
- ↑ Luo, Zhe-Xi; Ji, Qiang; Wible, John R.; Yuan, Chong-Xi (2003-12-12). "An early Cretaceous tribosphenic mammal and metatherian evolution". Science 302 (5652): 1934–1940. Bibcode:2003Sci...302.1934L. doi:10.1126/science.1090718. PMID 14671295. Retrieved 2010-12-27.
- ↑ Nilsson, M. A.; Churakov, G.; Sommer, M.; Van Tran, N.; Zemann, A.; Brosius, J.; Schmitz, J. (2010-07-27). Penny, David, ed. "Tracking Marsupial Evolution Using Archaic Genomic Retroposon Insertions". PLoS Biology (Public Library of Science) 8 (7): e1000436. doi:10.1371/journal.pbio.1000436. PMC 2910653. PMID 20668664.
- ↑ Woodburne, Michael O.; Zinsmeister, William J. (Oct 1982). "Fossil Land Mammal from Antarctica". Science 218 (4569): 284–286. Bibcode:1982Sci...218..284W. doi:10.1126/science.218.4569.284. PMID 17838631. Retrieved 2009-01-17.
- ↑ Goin, Francisco J.; et al. (Dec 1999). "New Discoveries of "Opposum-Like" Marsupials from Antarctica (Seymour Island, Medial Eocene)". Journal of Mammalian Evolution 6 (4): 335–365. doi:10.1023/A:1027357927460. Retrieved 2009-01-17.
- ↑ Reguero, Marcelo A.; Marenssi, Sergio A.; Santillana, Sergio N. (May 2002). "Antarctic Peninsula and South America (Patagonia) Paleogene terrestrial faunas and environments: biogeographic relationships". Palaeogeography, Palaeoclimatology, Palaeoecology 179 (3–4): 189–210. doi:10.1016/S0031-0182(01)00417-5.
- ↑ Mills, William James. Exploring Polar Frontiers: A Historical Encyclopedia, ABC-CLIO, 2003. ISBN 1-57607-422-6, ISBN 978-1-57607-422-0
- ↑ Goin, F.J.; Reguero, M.A.; Pascual, R.; von Koenigswald, W.; Woodburne, M.O.; Case, J.A.; Marenssi, S.A.; Vieytes, C.; Vizcaíno, S.F. (2006). "First gondwanatherian mammal from Antarctica". Geological Society, London, Special Publications 258: 135–144. Bibcode:2006GSLSP.258..135G. doi:10.1144/GSL.SP.2006.258.01.10.
- ↑ Prothero, Donald R.; Schoch, Robert M. (2002). Horns, tusks, and flippers: the evolution of hoofed mammals. JHU press. p. 45. ISBN 0-8018-7135-2.
- ↑ Tanaka, T.; Hashimoto, H. (1989). "Drug-resistance and its transferability of Shigella strains isolated in 1986 in Japan". Kansenshogaku Zasshi 63 (1): 15–26. PMID 2501419.
- ↑ Raven, Peter H. (2005). Biology of Plants (7th rev. ed.). New York: W.H. Freeman. ISBN 0-7167-6284-6. OCLC 183148564.
- ↑ Haas, J. W., Jr. (June 2000). "The Rev. Dr. William H. Dallinger F.R.S.: Early Advocate of Theistic Evolution and Foe of Spontaneous Generation". Perspectives on Science and Christian Faith 52: 107–117. Retrieved 2010-06-15.
- 1 2 Le Page, Michael (16 April 2008). "NS:bacteria make major evolutionary shift in the lab". New Scientist. Retrieved 9 July 2012.
- ↑ Blount ZD, Borland CZ, Lenski RE (June 2008). "Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli". Proceedings of the National Academy of Sciences of the United States of America 105 (23): 7899–906. Bibcode:2008PNAS..105.7899B. doi:10.1073/pnas.0803151105. PMC 2430337. PMID 18524956.
- ↑ Richard E. Lenski and Michael Travisano (1994). "Dynamics of adaptation and diversification: A 10,000-generation experiment with bacterial populations". Proc. Natl. Acad. Sci. 91: 6808–6814. Bibcode:1994PNAS...91.6808L. doi:10.1073/pnas.91.15.6808. PMC 44287. PMID 8041701. Retrieved 2015-03-17.
- 1 2 John A. Endler (1980). "Natural Selection on Color Patterns in Poecilia Reticulata". Evolution 34: 76–91. doi:10.2307/2408316.
- ↑ David N. Reznick, Frank H. Shaw, F. Helen Rodd and Ruth G. Shaw (1997). "Evaluation of the Rate of Evolution in Natural Populations of Guppies (Poecilia reticulata)". Science 275: 1934–1937. doi:10.1126/science.275.5308.1934. PMID 9072971.
- ↑ David Reznick and John A. Endler (1982). "The Impact of Predation on Life History Evolution in Trinidadian Guppies (Poecilia reticulata)". Evolution 36 (1): 160–177. doi:10.2307/2407978. JSTOR 2407978.
- ↑ Medical Research Council (UK) (November 21, 2009). "Brain Disease 'Resistance Gene' Evolves in Papua New Guinea Community; Could Offer Insights Into CJD". Science Daily (online) (Science News). Retrieved 2009-11-22.
- ↑ Mead, S.; Whitfield, J.; Poulter, M.; Shah, P.; Uphill, J.; Campbell, T.; Al-Dujaily, H.; Hummerich, H.; Beck, J.; Mein, C. A.; Verzilli, C.; Whittaker, J.; Alpers, M. P.; Collinge, J. (2009). "A Novel Protective Prion Protein Variant that Colocalizes with Kuru Exposure". The New England Journal of Medicine 361 (21): 2056–2065. doi:10.1056/NEJMoa0809716. PMID 19923577.
- ↑ Byars, S. G.; Ewbank, D.; Govindaraju, D. R.; Stearns, S. C. (2009). "Natural selection in a contemporary human population". Proceedings of the National Academy of Sciences 107 (suppl_1): 1787–1792. Bibcode:2010PNAS..107.1787B. doi:10.1073/pnas.0906199106. PMC 2868295. PMID 19858476.
- ↑ Soy and Lactose Intolerance Wayback: Soy Nutrition
- ↑ Enattah NS, Sahi T, Savilahti E, Terwilliger JD, Peltonen L, Järvelä I (2002). "Identification of a variant associated with adult-type hypolactasia". Nature Genetics 30 (2): 233–7. doi:10.1038/ng826. PMID 11788828.
- ↑ Curry, Andrew (31 July 2013). "Archaeology: The milk revolution". Nature.
- ↑ Swallow DM (2003). "Genetics of lactase persistence and lactose intolerance". Annual Review of Genetics 37: 197–219. doi:10.1146/annurev.genet.37.110801.143820. PMID 14616060.
- ↑ Harriet, Coles (2007-01-20). "The lactase gene in Africa: Do you take milk?". The Human Genome, Wellcome Trust. Retrieved 2008-07-18.
- ↑ Tishkoff SA, Reed FA, Ranciaro A; et al. (January 2007). "Convergent adaptation of human lactase persistence in Africa and Europe". Nature Genetics 39 (1): 31–40. doi:10.1038/ng1946. PMC 2672153. PMID 17159977.
- ↑ Welsh, Jennifer (February 17, 2011). "Fish Evolved to Survive GE Toxins in Hudson River". LiveScience. Retrieved 2011-02-19.
- 1 2 3 Isaac Wirgin; et al. (2011). "Mechanistic Basis of Resistance to PCBs in Atlantic Tomcod from the Hudson River". Science 331: 1322–1325. Bibcode:2011Sci...331.1322W. doi:10.1126/science.1197296.
- ↑ Clark, Bryan W.; et al. (2013). "Compound- and Mixture-Specific Differences in Resistance to Polycyclic Aromatic Hydrocarbons and PCB-126 among Fundulus heteroclitus Subpopulations throughout the Elizabeth River Estuary". Environmental Science & Technology 47: 10556–10566. doi:10.1021/es401604b.
- 1 2 Science News, Dark Power: Pigment seems to put radiation to good use, Week of May 26, 2007; Vol. 171, No. 21, p. 325 by Davide Castelvecchi
- ↑ Dadachova, E.; Bryan, R.A.; Huang, X.; Moadel, T.; Schweitzer, A.D.; Aisen, P.; Nosanchuk, J.D.; Casadevall, A. (2007). Rutherford, Julian, ed. "Ionizing Radiation Changes the Electronic Properties of Melanin and Enhances the Growth of Melanized Fungi". PLOS ONE 2 (5): e457. Bibcode:2007PLoSO...2..457D. doi:10.1371/journal.pone.0000457. PMC 1866175. PMID 17520016.
- ↑ Thwaites, W.M. (Summer 1985). "New Proteins Without God's Help". Creation Evolution Journal (National Center for Science Education (NCSE)) 5 (2): 1–3.
- ↑ Evolution and Information: The Nylon Bug. Nmsr.org. Retrieved on 2011-12-06.
- ↑ Why scientists dismiss 'intelligent design', Ker Than, MSNBC, Sept. 23, 2005
- ↑ Miller, Kenneth R. Only a Theory: Evolution and the Battle for America's Soul (2008) pp. 80–82
- ↑ Cheptou, P.; Carrue, O.; Rouifed, S.; Cantarel, A. (2008). "Rapid evolution of seed dispersal in an urban environment in the weed Crepis sancta". Proceedings of the National Academy of Sciences 105 (10): 3796–9. Bibcode:2008PNAS..105.3796C. doi:10.1073/pnas.0708446105. PMC 2268839. PMID 18316722.
- ↑ "Evolution in the urban jungle". Retrieved 2010-07-08.
- ↑ "Human Activity Boosts Brain Size in Animals". Yale Scientific. Retrieved 2015-03-16.
- ↑ Emilie C. Snell-Rood and Naomi Wick (2013). "Anthropogenic environments exert variable selection on cranial capacity in mammals" (PDF). R Soc B 280: 20131384. doi:10.1098/rspb.2013.1384. Retrieved 2015-03-16.
- 1 2 3 4 5 6 7 8 9 Herrel A, Huyghe K, Vanhooydonck B; et al. (March 2008). "Rapid large-scale evolutionary divergence in morphology and performance associated with exploitation of a different dietary resource". Proc. Natl. Acad. Sci. U.S.A. 105 (12): 4792–5. Bibcode:2008PNAS..105.4792H. doi:10.1073/pnas.0711998105. PMC 2290806. PMID 18344323.
- 1 2 3 4 Bart Vervust, Irena Grbac, Raoul Van Damme (August 2007). "Differences in morphology, performance and behaviour between recently diverged populations of Podarcis sicula mirror differences in predation pressure". Oikos 116 (8): 1343–1352. doi:10.1111/j.0030-1299.2007.15989.x.
- ↑ "Lizards Rapidly Evolve After Introduction to Island". National Geographic.
- ↑ Myers, PZ (23 April 2008). "Still just a lizard". ScienceBlogs.
- ↑ Sean Stankowski and Matthew A. Streisfeld (2015), "Introgressive hybridization facilitates adaptive divergence in a recent radiation of monkeyflowers", Proc. R. Soc. B 282: 20151666, doi:10.1098/rspb.2015.1666
- ↑ James K. Liebherr and Joseph V. McHugh in Resh, V. H. & R. T. Cardé (Editors) 2003. Encyclopedia of Insects. Academic Press.
- ↑ Bearhop, S.; Fiedler, W.; Furness, R.W.; Votier, S.C.; Waldron, S.; Newton, J.; Bowen, G.J.; Berthold, P.; Farnsworth, K. (2005). "Assortative mating as a mechanism for rapid evolution of a migratory divide". Science 310 (5747): 502–504. Bibcode:2005Sci...310..502B. doi:10.1126/science.1115661. PMID 16239479. Supporting Online Material
- ↑ Yong, Ed (December 3, 2009). "British birdfeeders split blackcaps into two genetically distinct groups : Not Exactly Rocket Science". ScienceBlogs. Retrieved 2010-05-21.
- ↑ Rice, William R.; Salt, George W. (1990). "The Evolution of Reproductive Isolation as a Correlated Character Under Sympatric Conditions: Experimental Evidence". Evolution, Society for the Study of Evolution 44.
- ↑ "The Evolution of Reproductive Isolation as a Correlated Character Under Sympatric Conditions: Experimental Evidence" (PDF). William R. Rice, George W. Salt. Archived from the original (PDF) on 2012-05-13. Retrieved 2010-05-23.
- ↑ "Observed Instances of Speciation, 5.3.5 Sympatric Speciation in Drosophila melanogaster". Joseph Boxhorn. Retrieved 2010-05-23.
- ↑ Scott P. Egan; et al. (2012), "Divergent host-plant use promotes reproductive isolation among cynipid gall wasp populations", Biology Letters 8: 605–608, doi:10.1098/rsbl.2011.1205
- ↑ Scott P. Egan; et al. (2013), "Parallel Patterns of Morphological and Behavioral Variation among Host-Associated Populations of Two Gall Wasp Species", PLOS ONE 8: e54690, Bibcode:2013PLoSO...854690E, doi:10.1371/journal.pone.0054690
- ↑ Feder, J.L.; Roethele, J.B.; Filchak, K.; Niedbalski, J.; Romero-Severson, J. (1 March 2003). "Evidence for inversion polymorphism related to sympatric host race formation in the apple maggot fly, Rhagoletis pomonella". Genetics 163 (3): 939–53. PMC 1462491. PMID 12663534.
- ↑ Berlocher, S.H.; Bush, G.L. (1982). "An electrophoretic analysis of Rhagoletis (Diptera: Tephritidae) phylogeny". Systematic Zoology 31 (2): 136–55. doi:10.2307/2413033. JSTOR 2413033.
- ↑ Berlocher, S.H.; Feder, J.L. (2002). "Sympatric speciation in phytophagous insects: moving beyond controversy?". Annu Rev Entomol. 47: 773–815. doi:10.1146/annurev.ento.47.091201.145312. PMID 11729091.
- ↑ Bush, G.L. (1969). "Sympatric host race formation and speciation in frugivorous flies of the genus Rhagoletis (Diptera: Tephritidae)". Evolution 23 (2): 237–51. doi:10.2307/2406788. JSTOR 2406788.
- ↑ Prokopy, R.J.; Diehl, S.R.; Cooley, S.S. (1988). "Behavioral evidence for host races in Rhagoletis pomonella flies". Oecologia 76 (1): 138–47. doi:10.1007/BF00379612. JSTOR 4218647.
- ↑ Feder, J.L.; Roethele, J.B.; Wlazlo, B.; Berlocher, S.H. (1997). "Selective maintenance of allozyme differences among sympatric host races of the apple maggot fly". Proc Natl Acad Sci USA. 94 (21): 11417–21. Bibcode:1997PNAS...9411417F. doi:10.1073/pnas.94.21.11417. PMC 23485. PMID 11038585.
- 1 2 3 "London underground source of new insect forms". The Times. 1998-08-26.
- 1 2 Fonseca, D. M.; Keyghobadi, N; Malcolm, CA; Mehmet, C; Schaffner, F; Mogi, M; Fleischer, RC; Wilkerson, RC (2004). "Emerging vectors in the Culex pipiens complex" (PDF). Science 303 (5663): 1535–8. Bibcode:2004Sci...303.1535F. doi:10.1126/science.1094247. PMID 15001783.
- 1 2 Burdick, Alan (2001). "Insect From the Underground — London, England Underground home to different species of mosquitos". Natural History.
- ↑ Byrne, K.; Nichols, R.A. (1999). "Culex pipiens in London Underground tunnels: differentiation between surface and subterranean populations". Heredity 82 (1): 7–15. doi:10.1038/sj.hdy.6884120. PMID 10200079.
- ↑ Vinogradova, E.B.; Shaikevich, E.V. (2007). "Morphometric, physiological and molecular characteristics of underground populations of the urban mosquito Culex pipiens Linnaeus f. molestus Forskål (Diptera: Culicidae) from several areas of Russia" (PDF). European Mosquito Bulletin 22: 17–24.
- ↑ Tobler, Michael (2009). "Does a predatory insect contribute to the divergence between cave- and surface-adapted fish populations?". Biology Letters 5: 506–509. doi:10.1098/rsbl.2009.0272.
- ↑ "Giant insect splits cavefish into distinct populations". Ed Yong. Retrieved 2010-05-22.
- ↑ J. P. Doupé, J. H. England, M. Furze and D. Paetkau (2007), "Most Northerly Observation of a Grizzly Bear (Ursus arctos) in Canada: Photographic and DNA Evidence from Melville Island, Northwest Territories", Artic 60: 271–276
- ↑ Gray, A. P. (1972), Mammalian hybrids. A check-list with bibliography (2nd ed.), Farnham Royal, Slough SL2 3BN, England: Commonwealth Agricultural Bureaux
- ↑ Kutschera, Verena E; Bidon, Tobias; Hailer, Frank; Rodi, Julia L.; Fain, Steven R.; Janke, Axel (2014). "Bears in a Forest of Gene Trees: Phylogenetic Inference Is Complicated by Incomplete Lineage Sorting and Gene Flow". Molecular Biology and Evolution 31 (8): 2004–2017. doi:10.1093/molbev/msu186.
- 1 2 Lindqvist, C.; Schuster, S. C.; Sun, Y.; Talbot, S. L.; Qi, J.; Ratan, A.; Tomsho, L. P.; Kasson, L.; Zeyl, E.; Aars, J.; Miller, W.; Ingolfsson, O.; Bachmann, L.; Wiig, O. (2010). "Complete mitochondrial genome of a Pleistocene jawbone unveils the origin of polar bear". Proceedings of the National Academy of Sciences 107 (11): 5053–7. Bibcode:2010PNAS..107.5053L. doi:10.1073/pnas.0914266107. PMC 2841953. PMID 20194737.
- ↑ Hailer, F.; Kutschera, V. E.; Hallstrom, B. M.; Klassert, D.; Fain, S. R.; Leonard, J. A.; Arnason, U.; Janke, A. (2012). "Nuclear Genomic Sequences Reveal that Polar Bears Are an Old and Distinct Bear Lineage". Science 336 (6079): 344–7. Bibcode:2012Sci...336..344H. doi:10.1126/science.1216424. PMID 22517859.
- ↑ Liu S.; et al. (2014), "Population genomics reveal recent speciation and rapid evolutionary adaptation in polar bears", Cell 157 (4): 785–794, doi:10.1016/j.cell.2014.03.054
- ↑ Miller, W; Schuster, SC; Welch, AJ; Ratan, A; Bedoya-Reina, OC; Zhao, F; Kim, HL; Burhans, RC; Drautz, DI; Wittekindt, NE; Tomsho, LP; Ibarra-Laclette, E; Herrera-Estrella, L; Peacock, E; Farley, S; Sage, GK; Rode, K; Obbard, M; Montiel, R; Bachmann, L; Ingólfsson, O; Aars, J; Mailund, T; Wiig, O; Talbot, SL; Lindqvist, C (2012). "Polar and brown bear genomes reveal ancient admixture and demographic footprints of past climate change". Proceedings of the National Academy of Sciences of the United States of America 109 (36): E2382–E2390. doi:10.1073/pnas.1210506109.
- ↑ "Evolution". Polar Bears International. Retrieved 30 January 2014.
- ↑ Hansman, Jared (6 August 2008). "Adaptive Traits of the Polar Bear (Ursus maritimus)". Helium, Inc. Retrieved 30 January 2014.
- 1 2 Bomblies, Kirsten; Lempe, Janne; Epple, Petra; Warthmann, Norman; Lanz, Christa; Dangl, Jeffery L.; Weigel, Detlef (2007). "Autoimmune Response as a Mechanism for a Dobzhansky-Muller-Type Incompatibility Syndrome in Plants". PLoS Biol 5 (9): e236. doi:10.1371/journal.pbio.0050236. PMC 1964774. PMID 17803357.
- ↑ "New plant species arise from conflicts between immune system genes". Ed Yong. Retrieved 2010-05-22.
- ↑ Paul N. Pearson, Thomas H. G. Ezard (2014). "Evolution and speciation in the Eocene planktonic foraminifer Turborotalia". Paleobiology 40: 130–143. doi:10.1666/13004.
- ↑ Vallejo-Marín, Mario (2012). "Mimulus peregrinus (Phrymaceae): A new British allopolyploid species". PhytoKeys 14: 1–14. doi:10.3897/phytokeys.14.3305. Retrieved 2014-01-27.
- ↑ Vallejo-Marín, Mario; Buggs, Richard J.; Cooley, Arielle M.; Puzey, Joshua R. (2015). "Speciation by genome duplication: Repeated origins and genomic composition of the recently formed allopolyploid species Mimulus peregrinus". Evolution 69: 1487–1500. doi:10.1111/evo.12678. Retrieved 2015-05-10.
- ↑ Karpechenko, G.D., Polyploid hybrids of Raphanus sativus X Brassica oleracea L., Bull. Appl. Bot. 17:305–408 (1927).
- ↑ Terasawa, Y. Crossing between Brassico-raphanus and B. chinensis and Raphanus sativus. Japanese Journal of Genetics. 8(4): 229–230 (1933).
- ↑ Lowe, Andrew J.; Abbott, Richard J. (1996). "Origins of the New Allopolyploid Species Senecio camrensis (asteracea) and its Relationship to the Canary Islands Endemic Senecio tenerifae". American Journal of Botany 83 (10): 1365–1372. doi:10.2307/2446125. JSTOR 2446125.
- ↑ Coyne, Jerry A. (2009). Why Evolution is True. Penguin Group. pp. 187–189. ISBN 978-0-670-02053-9.
- ↑ Missouri Botanical Garden. "TROPICOS Web display Senecio vulgaris L". Nomenclatural and Specimen Data Base. Missouri State Library. Retrieved 2008-02-01.
- ↑ Purves, William Kirkwood; Sadava, David E.; Orians, Gordon H.; Heller, H. Craig (2006). Life, the science of biology (7 ed.). Sinaur Associates, Inc. p. 487. ISBN 0-7167-9856-5.
- ↑ Soltis, Pam (2011-03-17). "UF researcher: Flowering plant study 'catches evolution in the act'". EurekAlert, American Association for the Advancement of Science. Retrieved 2011-03-28.
- ↑ Buggs, Richard J.A.; Zhang, Linjing; Miles, Nicholas; Tate, Jennifer A.; Gao, Lu; Wei, Wu; Schnable, Patrick S.; Barbazuk, W. Brad; Soltis, Pamela S. (2011). "Transcriptomic Shock Generates Evolutionary Novelty in a Newly Formed, Natural Allopolyploid Plant". Current Biology 21 (7): 551–6. doi:10.1016/j.cub.2011.02.016. PMID 21419627.
- ↑ Simulated Evolution Gets Complex. Trnmag.com (2003-05-08). Retrieved on 2011-12-06.
- ↑ Adami, C.; Ofria, C.; Collier, T.C. (2000). "Evolution of biological complexity". Proc Natl Acad Sci USA. 97 (9): 4463–8. arXiv:physics/0005074. Bibcode:2000PNAS...97.4463A. doi:10.1073/pnas.97.9.4463. PMC 18257. PMID 10781045.
- ↑ Earl, D.J.; Deem, M.W. (2004). "Evolvability is a selectable trait". Proc Natl Acad Sci USA. 101 (32): 11531–6. arXiv:q-bio/0407012. Bibcode:2004PNAS..10111531E. doi:10.1073/pnas.0404656101. PMC 511006. PMID 15289608.
- 1 2 Stemmer, W.P. (1994). "DNA shuffling by random fragmentation and reassembly: in vitro recombination for molecular evolution". Proc Natl Acad Sci USA. 91 (22): 10747–51. Bibcode:1994PNAS...9110747S. doi:10.1073/pnas.91.22.10747. PMC 45099. PMID 7938023.
- ↑ Sauter, E. (March 27, 2006). ""Accelerated Evolution" Converts RNA Enzyme to DNA Enzyme In Vitro". TSRI – News & Views 6 (11).
- ↑ Molecular evolution. kaist.ac.kr
- ↑ In Vitro Molecular Evolution. Isgec.org (1975-08-04). Retrieved on 2011-12-06.
- ↑ "Digital organisms used to confirm evolutionary process". American Association for the Advancement of Science. Retrieved 2011-03-21.
- ↑ "Artificial life experiments show how complex functions can evolve". American Association for the Advancement of Science. Retrieved 2011-03-21.
- ↑ Lenski, Richard E.; Ofria, Charles; Wilke, Claus O.; Wang, Jia Lan; Adami, Christoph (2001-07-19). "Evolution of digital organisms at high mutation rates leads to survival of the flattest". Nature 412 (6844): 331–3. Bibcode:2001Natur.412..331W. doi:10.1038/35085569. PMID 11460163.
{kind=link}
Further reading
- Biological science, Oxford, 2002.
- Clegg, C.J. (1998). Genetics & evolution. London: J. Murray. ISBN 0-7195-7552-4.
- Coyne, Jerry A. (2009). Why Evolution is True. New York: Oxford University Press. ISBN 978-0-19-923084-6.
- Darwin, Charles November 24, 1859. On the Origin of Species by means of Natural Selection or the Preservation of Favoured Races in the Struggle for Life. London: John Murray, Albemarle Street. 502 pages. Reprinted: Gramercy (May 22, 1995). ISBN 0-517-12320-7
- Peter R. Grant & B. Rosemary Grant (2014-04-06). 40 Years of Evolution: Darwin's Finches on Daphne Major Island. Princeton University Press. p. 432. ISBN 978-0691160467.
- Dawkins, Richard (2009). The Greatest Show on Earth: The Evidence for Evolution. Bantam Press. ISBN 978-1-4165-9478-9.
- Endler, John A. (1986). Natural selection in the wild. New Jersey: Princeton University Press. ISBN 0-691-08387-8.
- Hill, A.; Behrensmeyer, A.K. (1980). Fossils in the making: vertebrate taphonomy and paleoecology. Chicago: University of Chicago Press. ISBN 0-226-04169-7.
- Mayr, Ernst (2001). What evolution is. New York: Basic Books. ISBN 0-465-04426-3.
- Paul, C.R.C.; Donovan, S.K. (1998). The adequacy of the fossil record. New York: John Wiley. ISBN 0-471-96988-5.
- Shubin, Neil (2008). Your Inner Fish:A Journey Into the 3.5 Billion-Year History of the Human Body. Random House, Inc. ISBN 978-0-375-42447-2.
- Sober, Elliott (2008). Evidence and Evolution: The logic behind the science. Cambridge University Press. ISBN 978-0-521-87188-4.
External links
- National Academies Evolution Resources
- TalkOrigins Archive – 29+ Evidences for Macroevolution: The Scientific Case for Common Descent
- TalkOrigins Archive – Transitional Vertebrate Fossils FAQ
- Understanding Evolution: Your one-stop source for information on evolution
- National Academy Press: Teaching About Evolution and the Nature of Science
- Evolution — Provided by PBS.
- Evolution News from Genome News Network (GNN)
- Evolution by Natural Selection — An introduction to the logic of the theory of evolution by natural selection
- Howstuffworks.com — How Evolution Works
- 15 Evolutionary Gems
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||