Enzyme kinetics

Enzyme kinetics is the study of the chemical reactions that are catalysed by enzymes. In enzyme kinetics, the reaction rate is measured and the effects of varying the conditions of the reaction are investigated. Studying an enzyme's kinetics in this way can reveal the catalytic mechanism of this enzyme, its role in metabolism, how its activity is controlled, and how a drug or an agonist might inhibit the enzyme.
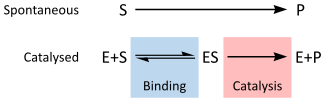
Enzymes are usually protein molecules that manipulate other molecules — the enzymes' substrates. These target molecules bind to an enzyme's active site and are transformed into products through a series of steps known as the enzymatic mechanism
E + S ⇄ ES ⇄ ES* ⇄ EP ⇄ E + P
These mechanisms can be divided into single-substrate and multiple-substrate mechanisms. Kinetic studies on enzymes that only bind one substrate, such as triosephosphate isomerase, aim to measure the affinity with which the enzyme binds this substrate and the turnover rate. Some other examples of enzymes are phosphofructokinase and hexokinase, both of which are important for cellular respiration (glycolysis).
When enzymes bind multiple substrates, such as dihydrofolate reductase (shown right), enzyme kinetics can also show the sequence in which these substrates bind and the sequence in which products are released. An example of enzymes that bind a single substrate and release multiple products are proteases, which cleave one protein substrate into two polypeptide products. Others join two substrates together, such as DNA polymerase linking a nucleotide to DNA. Although these mechanisms are often a complex series of steps, there is typically one rate-determining step that determines the overall kinetics. This rate-determining step may be a chemical reaction or a conformational change of the enzyme or substrates, such as those involved in the release of product(s) from the enzyme.
Knowledge of the enzyme's structure is helpful in interpreting kinetic data. For example, the structure can suggest how substrates and products bind during catalysis; what changes occur during the reaction; and even the role of particular amino acid residues in the mechanism. Some enzymes change shape significantly during the mechanism; in such cases, it is helpful to determine the enzyme structure with and without bound substrate analogues that do not undergo the enzymatic reaction.
Not all biological catalysts are protein enzymes; RNA-based catalysts such as ribozymes and ribosomes are essential to many cellular functions, such as RNA splicing and translation. The main difference between ribozymes and enzymes is that RNA catalysts are composed of nucleotides, whereas enzymes are composed of amino acids. Ribozymes also perform a more limited set of reactions, although their reaction mechanisms and kinetics can be analysed and classified by the same methods.
General principles
.svg.png)
 . Beyond this limit the enzyme is saturated with substrate and the reaction rate ceases to increase.
. Beyond this limit the enzyme is saturated with substrate and the reaction rate ceases to increase.The reaction catalysed by an enzyme uses exactly the same reactants and produces exactly the same products as the uncatalysed reaction. Like other catalysts, enzymes do not alter the position of equilibrium between substrates and products.[1] However, unlike uncatalysed chemical reactions, enzyme-catalysed reactions display saturation kinetics. For a given enzyme concentration and for relatively low substrate concentrations, the reaction rate increases linearly with substrate concentration; the enzyme molecules are largely free to catalyse the reaction, and increasing substrate concentration means an increasing rate at which the enzyme and substrate molecules encounter one another. However, at relatively high substrate concentrations, the reaction rate asymptotically approaches the theoretical maximum; the enzyme active sites are almost all occupied and the reaction rate is determined by the intrinsic turnover rate of the enzyme. The substrate concentration midway between these two limiting cases is denoted by KM.
The two most important kinetic properties of an enzyme are how quickly the enzyme becomes saturated with a particular substrate, and the maximum rate it can achieve. Knowing these properties suggests what an enzyme might do in the cell and can show how the enzyme will respond to changes in these conditions.
Enzyme assays
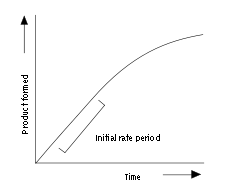
Enzyme assays are laboratory procedures that measure the rate of enzyme reactions. Because enzymes are not consumed by the reactions they catalyse, enzyme assays usually follow changes in the concentration of either substrates or products to measure the rate of reaction. There are many methods of measurement. Spectrophotometric assays observe change in the absorbance of light between products and reactants; radiometric assays involve the incorporation or release of radioactivity to measure the amount of product made over time. Spectrophotometric assays are most convenient since they allow the rate of the reaction to be measured continuously. Although radiometric assays require the removal and counting of samples (i.e., they are discontinuous assays) they are usually extremely sensitive and can measure very low levels of enzyme activity.[2] An analogous approach is to use mass spectrometry to monitor the incorporation or release of stable isotopes as substrate is converted into product.
The most sensitive enzyme assays use lasers focused through a microscope to observe changes in single enzyme molecules as they catalyse their reactions. These measurements either use changes in the fluorescence of cofactors during an enzyme's reaction mechanism, or of fluorescent dyes added onto specific sites of the protein to report movements that occur during catalysis.[3] These studies are providing a new view of the kinetics and dynamics of single enzymes, as opposed to traditional enzyme kinetics, which observes the average behaviour of populations of millions of enzyme molecules.[4][5]
An example progress curve for an enzyme assay is shown above. The enzyme produces product at an initial rate that is approximately linear for a short period after the start of the reaction. As the reaction proceeds and substrate is consumed, the rate continuously slows (so long as substrate is not still at saturating levels). To measure the initial (and maximal) rate, enzyme assays are typically carried out while the reaction has progressed only a few percent towards total completion. The length of the initial rate period depends on the assay conditions and can range from milliseconds to hours. However, equipment for rapidly mixing liquids allows fast kinetic measurements on initial rates of less than one second.[6] These very rapid assays are essential for measuring pre-steady-state kinetics, which are discussed below.
Most enzyme kinetics studies concentrate on this initial, approximately linear part of enzyme reactions. However, it is also possible to measure the complete reaction curve and fit this data to a non-linear rate equation. This way of measuring enzyme reactions is called progress-curve analysis.[7] This approach is useful as an alternative to rapid kinetics when the initial rate is too fast to measure accurately.
Single-substrate reactions
Enzymes with single-substrate mechanisms include isomerases such as triosephosphateisomerase or bisphosphoglycerate mutase, intramolecular lyases such as adenylate cyclase and the hammerhead ribozyme, an RNA lyase.[8] However, some enzymes that only have a single substrate do not fall into this category of mechanisms. Catalase is an example of this, as the enzyme reacts with a first molecule of hydrogen peroxide substrate, becomes oxidised and is then reduced by a second molecule of substrate. Although a single substrate is involved, the existence of a modified enzyme intermediate means that the mechanism of catalase is actually a ping–pong mechanism, a type of mechanism that is discussed in the Multi-substrate reactions section below.
Michaelis–Menten kinetics

As enzyme-catalysed reactions are saturable, their rate of catalysis does not show a linear response to increasing substrate. If the initial rate of the reaction is measured over a range of substrate concentrations (denoted as [S]), the reaction rate (v) increases as [S] increases, as shown on the right. However, as [S] gets higher, the enzyme becomes saturated with substrate and the rate reaches Vmax, the enzyme's maximum rate.
The Michaelis–Menten kinetic model of a single-substrate reaction is shown on the right. There is an initial bimolecular reaction between the enzyme E and substrate S to form the enzyme–substrate complex ES but the rate of enzymatic reaction increases with the increase of the substrate concentration up to a certain level but then an increase in substrate concentration does not cause any increase in reaction rate as there no more E available for reacting with S and the rate of reaction becomes dependent on ES and the reaction becomes unimolecular reaction. Although the enzymatic mechanism for the unimolecular reaction 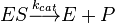 can be quite complex, there is typically one rate-determining enzymatic step that allows this reaction to be modelled as a single catalytic step with an apparent unimolecular rate constant kcat.
If the reaction path proceeds over one or several intermediates, kcat will be a function of several elementary rate constants, whereas in the simplest case of a single elementary reaction (e.g. no intermediates) it will be identical to the elementary unimolecular rate constant k2. The apparent unimolecular rate constant kcat is also called turnover number and denotes the maximum number of enzymatic reactions catalysed per second.
can be quite complex, there is typically one rate-determining enzymatic step that allows this reaction to be modelled as a single catalytic step with an apparent unimolecular rate constant kcat.
If the reaction path proceeds over one or several intermediates, kcat will be a function of several elementary rate constants, whereas in the simplest case of a single elementary reaction (e.g. no intermediates) it will be identical to the elementary unimolecular rate constant k2. The apparent unimolecular rate constant kcat is also called turnover number and denotes the maximum number of enzymatic reactions catalysed per second.
The Michaelis–Menten equation[9] describes how the (initial) reaction rate v0 depends on the position of the substrate-binding equilibrium and the rate constant k2.
![v_0 = \frac{V_{\max}[\mbox{S}]}{K_M + [\mbox{S}]}](../I/m/3429bb01497e614af0b197fe2c2979ca.png) (Michaelis–Menten equation)
(Michaelis–Menten equation)
with the constants
![\begin{align}
K_M \ &\stackrel{\mathrm{def}}{=}\ \frac{k_{2} + k_{-1}}{k_{1}} \approx K_D\\
V_\max \ &\stackrel{\mathrm{def}}{=}\ k_{cat}{[}E{]}_{tot}
\end{align}](../I/m/7cc86c5f51e917dfb3b517c937089935.png)
This Michaelis–Menten equation is the basis for most single-substrate enzyme kinetics. Two crucial assumptions underlie this equation (apart from the general assumption about the mechanism only involving no intermediate or product inhibition, and there is no allostericity or cooperativity). The first assumption is the so-called
quasi-steady-state assumption (or pseudo-steady-state hypothesis), namely that the concentration of the substrate-bound enzyme (and hence also the unbound enzyme) changes much more slowly than those of the product and substrate and thus the change over time of the complex can be set to zero
![d{[}ES{]}/{dt} \; \overset{!} = \;0](../I/m/08f59e2b77fc9b255da74af77b16dc0a.png) . The second assumption is that the total enzyme concentration does not change over time, thus
. The second assumption is that the total enzyme concentration does not change over time, thus ![{[}E{]}_\text{tot} = {[}E{]} + {[}ES{]} \; \overset{!} = \; \text{const}](../I/m/4667651f6f6d82a0c5bef9130f3af073.png) .
A complete derivation can be found here.
.
A complete derivation can be found here.
The Michaelis constant KM is experimentally defined as the concentration at which the rate of the enzyme reaction is half Vmax, which can be verified by substituting [S] = Km into the Michaelis–Menten equation and can also be seen graphically. If the rate-determining enzymatic step is slow compared to substrate dissociation (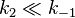 ), the Michaelis constant KM is roughly the dissociation constant KD of the ES complex.
), the Michaelis constant KM is roughly the dissociation constant KD of the ES complex.
If ![[S]](../I/m/38bd6c19740e43396bd14b1575d58f60.png) is small compared to
is small compared to 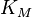 then the term
then the term ![[S] / (K_M + [S]) \approx [S] / K_M](../I/m/7071e21a580398420d8e6bd0673ea3df.png) and also very little ES complex is formed, thus
and also very little ES complex is formed, thus ![[E]_0 \approx [E]](../I/m/0f92ba855b6682e83c7195e3899887be.png) . Therefore, the rate of product formation is
. Therefore, the rate of product formation is
![v_0 \approx \frac{k_{cat}}{K_M} [E] [S] \qquad \qquad \text{if } [S] \ll K_M](../I/m/87214feb1ac3e7a5919b4e60a9ed3ddb.png)
Thus the product formation rate depends on the enzyme concentration as well as on the substrate concentration, the equation resembles a bimolecular reaction with a corresponding pseudo-second order rate constant 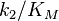 . This constant is a measure of catalytic efficiency. The most efficient enzymes reach a in the range of 108 – 1010 M−1 s−1. These enzymes are so efficient they effectively catalyse a reaction each time they encounter a substrate molecule and have thus reached an upper theoretical limit for efficiency (diffusion limit); and are sometimes referred to as kinetically perfect enzymes.[10]
. This constant is a measure of catalytic efficiency. The most efficient enzymes reach a in the range of 108 – 1010 M−1 s−1. These enzymes are so efficient they effectively catalyse a reaction each time they encounter a substrate molecule and have thus reached an upper theoretical limit for efficiency (diffusion limit); and are sometimes referred to as kinetically perfect enzymes.[10]
Direct use of the Michaelis–Menten equation for time course kinetic analysis
The observed velocities predicted by the Michaelis–Menten equation can be used to directly model the time course disappearance of substrate and the production of product through incorporation of the Michaelis–Menten equation into the equation for first order chemical kinetics. This can only be achieved however if one recognises the problem associated with the use of Euler's number in the description of first order chemical kinetics. i.e. e−k is a split constant that introduces a systematic error into calculations and can be rewritten as a single constant which represents the remaining substrate after each time period.[11]
![[S]=[S]_0(1-k)^{t}\,](../I/m/0cf28d29fb778a9f0c8c0e7c9962e8ec.png)
![[S]=[S]_0(1-v/[S]_0)^{t}\,](../I/m/52c183bd31bb0b670a6b5cd66c0b059e.png)
![[S]=[S]_0(1-(V_{\max} [S]_0 / (K_M + [S]_0)/[S]_0))^{t}\,](../I/m/3cf1684ecaa7f5b2ed4edd2fdbc17050.png)
In 1983 Stuart Beal (and also independently Santiago Schnell and Claudio Mendoza in 1997) derived a closed form solution for the time course kinetics analysis of the Michaelis-Menten mechanism.[12][13] The solution, known as the Schnell-Mendoza equation, has the form:
![\frac{[S]}{K_M} = W \left[ F(t) \right]\,](../I/m/37912a822dfc9a868126d0d23af27663.png)
where W[] is the Lambert-W function.[14][15] and where F(t) is
![F(t) = \frac{[S]_0}{K_M} \exp\!\left(\frac{[S]_0}{K_M} - \frac{V_\max}{K_M}\,t \right) \,](../I/m/ba4c7db00b78082b944a8a85c9d1ec4e.png)
This equation is encompassed by the equation below, obtained by Berberan-Santos (MATCH Commun. Math. Comput. Chem. 63 (2010) 283), which is also valid when the initial substrate concentration is close to that of enzyme,
![\frac{[S]}{K_M} = W \left[ F(t) \right]- \frac{V_\max}{k_{cat} K_M}\ \frac{W \left[ F(t) \right]}{1+W \left[ F(t) \right]}\,](../I/m/625c84ddb48f10df15140fc1a8142e8f.png)
where W[] is again the Lambert-W function.
Linear plots of the Michaelis–Menten equation

The plot of v versus [S] above is not linear; although initially linear at low [S], it bends over to saturate at high [S]. Before the modern era of nonlinear curve-fitting on computers, this nonlinearity could make it difficult to estimate KM and Vmax accurately. Therefore, several researchers developed linearisations of the Michaelis–Menten equation, such as the Lineweaver–Burk plot, the Eadie–Hofstee diagram and the Hanes–Woolf plot. All of these linear representations can be useful for visualising data, but none should be used to determine kinetic parameters, as computer software is readily available that allows for more accurate determination by nonlinear regression methods.[16]
The Lineweaver–Burk plot or double reciprocal plot is a common way of illustrating kinetic data. This is produced by taking the reciprocal of both sides of the Michaelis–Menten equation. As shown on the right, this is a linear form of the Michaelis–Menten equation and produces a straight line with the equation y = mx + c with a y-intercept equivalent to 1/Vmax and an x-intercept of the graph representing −1/KM.
![\frac{1}{v} = \frac{K_{M}}{V_{\max} [\mbox{S}]} + \frac{1}{V_\max}](../I/m/9fe6d77cc0fa7fe2cdb516e63c622f71.png)
Naturally, no experimental values can be taken at negative 1/[S]; the lower limiting value 1/[S] = 0 (the y-intercept) corresponds to an infinite substrate concentration, where 1/v=1/Vmax as shown at the right; thus, the x-intercept is an extrapolation of the experimental data taken at positive concentrations. More generally, the Lineweaver–Burk plot skews the importance of measurements taken at low substrate concentrations and, thus, can yield inaccurate estimates of Vmax and KM.[17] A more accurate linear plotting method is the Eadie-Hofstee plot. In this case, v is plotted against v/[S]. In the third common linear representation, the Hanes-Woolf plot, [S]/v is plotted against [S]. In general, data normalisation can help diminish the amount of experimental work and can increase the reliability of the output, and is suitable for both graphical and numerical analysis.[18]
Practical significance of kinetic constants
The study of enzyme kinetics is important for two basic reasons. Firstly, it helps explain how enzymes work, and secondly, it helps predict how enzymes behave in living organisms. The kinetic constants defined above, KM and Vmax, are critical to attempts to understand how enzymes work together to control metabolism.
Making these predictions is not trivial, even for simple systems. For example, oxaloacetate is formed by malate dehydrogenase within the mitochondrion. Oxaloacetate can then be consumed by citrate synthase, phosphoenolpyruvate carboxykinase or aspartate aminotransferase, feeding into the citric acid cycle, gluconeogenesis or aspartic acid biosynthesis, respectively. Being able to predict how much oxaloacetate goes into which pathway requires knowledge of the concentration of oxaloacetate as well as the concentration and kinetics of each of these enzymes. This aim of predicting the behaviour of metabolic pathways reaches its most complex expression in the synthesis of huge amounts of kinetic and gene expression data into mathematical models of entire organisms. Alternatively, one useful simplification of the metabolic modelling problem is to ignore the underlying enzyme kinetics and only rely on information about the reaction network's stoichiometry, a technique called flux balance analysis.[19][20]
Michaelis–Menten kinetics with intermediate
One could also consider the less simple case
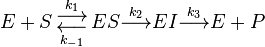
where a complex with the enzyme and an intermediate exists and the intermediate is converted into product in a second step. In this case we have a very similar equation[21]
![\begin{align}
v_0 &= k_{cat}\frac{{[}S{]} {[}E{]}_0}{K_M^{\prime}+ {[}S{]}}
\end{align}](../I/m/609a1bff6d5254c7a6aa0330a5f5a0b4.png)
but the constants are different

We see that for the limiting case 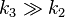 , thus when the last step from EI to E + P is much faster than the previous step, we get again the original equation. Mathematically we have then
, thus when the last step from EI to E + P is much faster than the previous step, we get again the original equation. Mathematically we have then 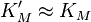 and
and  .
.
Multi-substrate reactions
Multi-substrate reactions follow complex rate equations that describe how the substrates bind and in what sequence. The analysis of these reactions is much simpler if the concentration of substrate A is kept constant and substrate B varied. Under these conditions, the enzyme behaves just like a single-substrate enzyme and a plot of v by [S] gives apparent KM and Vmax constants for substrate B. If a set of these measurements is performed at different fixed concentrations of A, these data can be used to work out what the mechanism of the reaction is. For an enzyme that takes two substrates A and B and turns them into two products P and Q, there are two types of mechanism: ternary complex and ping–pong.
Ternary-complex mechanisms

In these enzymes, both substrates bind to the enzyme at the same time to produce an EAB ternary complex. The order of binding can either be random (in a random mechanism) or substrates have to bind in a particular sequence (in an ordered mechanism). When a set of v by [S] curves (fixed A, varying B) from an enzyme with a ternary-complex mechanism are plotted in a Lineweaver–Burk plot, the set of lines produced will intersect.
Enzymes with ternary-complex mechanisms include glutathione S-transferase,[22] dihydrofolate reductase[23] and DNA polymerase.[24] The following links show short animations of the ternary-complex mechanisms of the enzymes dihydrofolate reductase[β] and DNA polymerase[γ].
Ping–pong mechanisms
As shown on the right, enzymes with a ping-pong mechanism can exist in two states, E and a chemically modified form of the enzyme E*; this modified enzyme is known as an intermediate. In such mechanisms, substrate A binds, changes the enzyme to E* by, for example, transferring a chemical group to the active site, and is then released. Only after the first substrate is released can substrate B bind and react with the modified enzyme, regenerating the unmodified E form. When a set of v by [S] curves (fixed A, varying B) from an enzyme with a ping–pong mechanism are plotted in a Lineweaver–Burk plot, a set of parallel lines will be produced. This is called a secondary plot.
Enzymes with ping–pong mechanisms include some oxidoreductases such as thioredoxin peroxidase,[25] transferases such as acylneuraminate cytidylyltransferase[26] and serine proteases such as trypsin and chymotrypsin.[27] Serine proteases are a very common and diverse family of enzymes, including digestive enzymes (trypsin, chymotrypsin, and elastase), several enzymes of the blood clotting cascade and many others. In these serine proteases, the E* intermediate is an acyl-enzyme species formed by the attack of an active site serine residue on a peptide bond in a protein substrate. A short animation showing the mechanism of chymotrypsin is linked here.[δ]
Non-Michaelis–Menten kinetics

Some enzymes produce a sigmoid v by [S] plot, which often indicates cooperative binding of substrate to the active site. This means that the binding of one substrate molecule affects the binding of subsequent substrate molecules. This behavior is most common in multimeric enzymes with several interacting active sites.[28] Here, the mechanism of cooperation is similar to that of hemoglobin, with binding of substrate to one active site altering the affinity of the other active sites for substrate molecules. Positive cooperativity occurs when binding of the first substrate molecule increases the affinity of the other active sites for substrate. Negative cooperativity occurs when binding of the first substrate decreases the affinity of the enzyme for other substrate molecules.
Allosteric enzymes include mammalian tyrosyl tRNA-synthetase, which shows negative cooperativity,[29] and bacterial aspartate transcarbamoylase[30] and phosphofructokinase,[31] which show positive cooperativity.
Cooperativity is surprisingly common and can help regulate the responses of enzymes to changes in the concentrations of their substrates. Positive cooperativity makes enzymes much more sensitive to [S] and their activities can show large changes over a narrow range of substrate concentration. Conversely, negative cooperativity makes enzymes insensitive to small changes in [S].
The Hill equation (biochemistry)[32] is often used to describe the degree of cooperativity quantitatively in non-Michaelis–Menten kinetics. The derived Hill coefficient n measures how much the binding of substrate to one active site affects the binding of substrate to the other active sites. A Hill coefficient of <1 indicates negative cooperativity and a coefficient of >1 indicates positive cooperativity.
Pre-steady-state kinetics

In the first moment after an enzyme is mixed with substrate, no product has been formed and no intermediates exist. The study of the next few milliseconds of the reaction is called Pre-steady-state kinetics also referred to as Burst kinetics. Pre-steady-state kinetics is therefore concerned with the formation and consumption of enzyme–substrate intermediates (such as ES or E*) until their steady-state concentrations are reached.
This approach was first applied to the hydrolysis reaction catalysed by chymotrypsin.[33] Often, the detection of an intermediate is a vital piece of evidence in investigations of what mechanism an enzyme follows. For example, in the ping–pong mechanisms that are shown above, rapid kinetic measurements can follow the release of product P and measure the formation of the modified enzyme intermediate E*.[34] In the case of chymotrypsin, this intermediate is formed by an attack on the substrate by the nucleophilic serine in the active site and the formation of the acyl-enzyme intermediate.
In the figure to the right, the enzyme produces E* rapidly in the first few seconds of the reaction. The rate then slows as steady state is reached. This rapid burst phase of the reaction measures a single turnover of the enzyme. Consequently, the amount of product released in this burst, shown as the intercept on the y-axis of the graph, also gives the amount of functional enzyme which is present in the assay.[35]
Chemical mechanism
An important goal of measuring enzyme kinetics is to determine the chemical mechanism of an enzyme reaction, i.e., the sequence of chemical steps that transform substrate into product. The kinetic approaches discussed above will show at what rates intermediates are formed and inter-converted, but they cannot identify exactly what these intermediates are.
Kinetic measurements taken under various solution conditions or on slightly modified enzymes or substrates often shed light on this chemical mechanism, as they reveal the rate-determining step or intermediates in the reaction. For example, the breaking of a covalent bond to a hydrogen atom is a common rate-determining step. Which of the possible hydrogen transfers is rate determining can be shown by measuring the kinetic effects of substituting each hydrogen by deuterium, its stable isotope. The rate will change when the critical hydrogen is replaced, due to a primary kinetic isotope effect, which occurs because bonds to deuterium are harder to break than bonds to hydrogen.[36] It is also possible to measure similar effects with other isotope substitutions, such as 13C/12C and 18O/16O, but these effects are more subtle.[37]
Isotopes can also be used to reveal the fate of various parts of the substrate molecules in the final products. For example, it is sometimes difficult to discern the origin of an oxygen atom in the final product; since it may have come from water or from part of the substrate. This may be determined by systematically substituting oxygen's stable isotope 18O into the various molecules that participate in the reaction and checking for the isotope in the product.[38] The chemical mechanism can also be elucidated by examining the kinetics and isotope effects under different pH conditions,[39] by altering the metal ions or other bound cofactors,[40] by site-directed mutagenesis of conserved amino acid residues, or by studying the behaviour of the enzyme in the presence of analogues of the substrate(s).[41]
Enzyme inhibition and activation

Enzyme inhibitors are molecules that reduce or abolish enzyme activity, while enzyme activators are molecules that increase the catalytic rate of enzymes. These interactions can be either reversible (i.e., removal of the inhibitor restores enzyme activity) or irreversible (i.e., the inhibitor permanently inactivates the enzyme).
Reversible inhibitors
Traditionally reversible enzyme inhibitors have been classified as competitive, uncompetitive, or non-competitive, according to their effects on Km and Vmax. These different effects result from the inhibitor binding to the enzyme E, to the enzyme–substrate complex ES, or to both, respectively. The division of these classes arises from a problem in their derivation and results in the need to use two different binding constants for one binding event. The binding of an inhibitor and its effect on the enzymatic activity are two distinctly different things, another problem the traditional equations fail to acknowledge. In noncompetitive inhibition the binding of the inhibitor results in 100% inhibition of the enzyme only, and fails to consider the possibility of anything in between.[42] The common form of the inhibitory term also obscures the relationship between the inhibitor binding to the enzyme and its relationship to any other binding term be it the Michaelis–Menten equation or a dose response curve associated with ligand receptor binding. To demonstrate the relationship the following rearrangement can be made:
![\cfrac{V_\max}{1 + \cfrac{[I]}{K_i}}](../I/m/cb29794646d4b42589b7599240e9df6c.png)
![\cfrac{V_\max}{\cfrac{[I]+K_i}{K_i}}](../I/m/cc2226956d00a276914a90c2fb127282.png)
Adding zero to the bottom ([I]-[I])
![\cfrac{V_\max}{\cfrac{[I]+K_i}{[I]+K_i-[I]}}](../I/m/a15ba82a47775ae7e5b143e001f11915.png)
Dividing by [I]+Ki
![\cfrac{V_\max}{\cfrac{1}{1 - \cfrac{[I]}{[I]+K_i}}}](../I/m/df7e09e16f571524a31438696f1ca00a.png)
![V_\max - V_\max \cfrac{[I]}{[I]+K_i}](../I/m/3f827c511f30ae697948fafd96714fc9.png)
This notation demonstrates that similar to the Michaelis–Menten equation, where the rate of reaction depends on the percent of the enzyme population interacting with substrate
fraction of the enzyme population bound by substrate
![\cfrac{[S]}{[S]+K_m}](../I/m/9a06a04189138ba95d32faceba858e59.png)
fraction of the enzyme population bound by inhibitor
![\cfrac{[I]}{[I]+K_i}](../I/m/2a6aad2f3c5e05e9fde4c6233b4494f0.png)
the effect of the inhibitor is a result of the percent of the enzyme population interacting with inhibitor. The only problem with this equation in its present form is that it assumes absolute inhibition of the enzyme with inhibitor binding, when in fact there can be a wide range of effects anywhere from 100% inhibition of substrate turn over to just >0%. To account for this the equation can be easily modified to allow for different degrees of inhibition by including a delta Vmax term.
![V_\max - \Delta V_\max \cfrac{[I]}{[I]+K_i}](../I/m/6d799921424129ce9d56d4369bdecd22.png)
or
![V_\max1 - (V_\max1 - V_\max2 ) \cfrac{[I]}{[I]+K_i}](../I/m/f28ee8573390c11378d08c860a79a761.png)
This term can then define the residual enzymatic activity present when the inhibitor is interacting with individual enzymes in the population. However the inclusion of this term has the added value of allowing for the possibility of activation if the secondary Vmax term turns out to be higher than the initial term. To account for the possibly of activation as well the notation can then be rewritten replacing the inhibitor "I" with a modifier term denoted here as "X".
![V_\max1 - (V_\max1 - V_\max2 ) \cfrac{[X]}{[X]+K_x}](../I/m/9c265a0a468de3f2d48392f712814879.png)
While this terminology results in a simplified way of dealing with kinetic effects relating to the maximum velocity of the Michaelis–Menten equation, it highlights potential problems with the term used to describe effects relating to the Km. The Km relating to the affinity of the enzyme for the substrate should in most cases relate to potential changes in the binding site of the enzyme which would directly result from enzyme inhibitor interactions. As such a term similar to the one proposed above to modulate Vmax should be appropriate in most situations:[43][44]
![K_m1 - (K_m1 - K_m2 ) \cfrac{[X]}{[X]+K_x}](../I/m/9ec46301608baef90a3f2c5514b25094.png)
Irreversible inhibitors
Enzyme inhibitors can also irreversibly inactivate enzymes, usually by covalently modifying active site residues. These reactions, which may be called suicide substrates, follow exponential decay functions and are usually saturable. Below saturation, they follow first order kinetics with respect to inhibitor.
Mechanisms of catalysis

The favoured model for the enzyme–substrate interaction is the induced fit model.[45] This model proposes that the initial interaction between enzyme and substrate is relatively weak, but that these weak interactions rapidly induce conformational changes in the enzyme that strengthen binding. These conformational changes also bring catalytic residues in the active site close to the chemical bonds in the substrate that will be altered in the reaction.[46] Conformational changes can be measured using circular dichroism or dual polarisation interferometry. After binding takes place, one or more mechanisms of catalysis lower the energy of the reaction's transition state by providing an alternative chemical pathway for the reaction. Mechanisms of catalysis include catalysis by bond strain; by proximity and orientation; by active-site proton donors or acceptors; covalent catalysis and quantum tunnelling.[34][47]
Enzyme kinetics cannot prove which modes of catalysis are used by an enzyme. However, some kinetic data can suggest possibilities to be examined by other techniques. For example, a ping–pong mechanism with burst-phase pre-steady-state kinetics would suggest covalent catalysis might be important in this enzyme's mechanism. Alternatively, the observation of a strong pH effect on Vmax but not Km might indicate that a residue in the active site needs to be in a particular ionisation state for catalysis to occur.
History
In 1902 Victor Henri proposed a quantitative theory of enzyme kinetics,[48] but at the time the experimental significance of the hydrogen ion concentration was not yet recognized. After Peter Lauritz Sørensen had defined the logarithmic pH-scale and introduced the concept of buffering in 1909[49] the German chemist Leonor Michaelis and his Canadian postdoc Maud Leonora Menten repeated Henri's experiments and confirmed his equation, which is now generally referred to as Michaelis-Menten kinetics (sometimes also Henri-Michaelis-Menten kinetics).[50] Their work was further developed by G. E. Briggs and J. B. S. Haldane, who derived kinetic equations that are still widely considered today a starting point in modeling enzymatic activity.[51]
The major contribution of the Henri-Michaelis-Menten approach was to think of enzyme reactions in two stages. In the first, the substrate binds reversibly to the enzyme, forming the enzyme-substrate complex. This is sometimes called the Michaelis complex. The enzyme then catalyzes the chemical step in the reaction and releases the product. The kinetics of many enzymes is adequately described by the simple Michaelis-Menten model, but all enzymes have internal motions that are not accounted for in the model and can have significant contributions to the overall reaction kinetics. This can be modeled by introducing several Michaelis-Menten pathways that are connected with fluctuating rates,[52][53][54] which is a mathematical extension of the basic Michaelis Menten mechanism.[55]
Software
ENZO
ENZO (Enzyme Kinetics) is a graphical interface tool for building kinetic models of enzyme catalyzed reactions. ENZO automatically generates the corresponding differential equations from a stipulated enzyme reaction scheme. These differential equations are processed by a numerical solver and a regression algorithm which fits the coefficients of differential equations to experimentally observed time course curves. ENZO allows rapid evaluation of rival reaction schemes and can be used for routine tests in enzyme kinetics.[56]
See also
Footnotes
α. ^ Link: Interactive Michaelis–Menten kinetics tutorial (Java required)
β. ^ Link: dihydrofolate reductase mechanism (Gif)
γ. ^ Link: DNA polymerase mechanism (Gif)
δ. ^ Link: Chymotrypsin mechanism (Flash required)
References
- ↑ Wrighton, Mark S.; Ebbing, Darrell D. (1993). General chemistry (4th ed.). Boston: Houghton Mifflin. ISBN 0-395-63696-5.
- ↑ Danson, Michael; Eisenthal, Robert (2002). Enzyme assays: a practical approach. Oxford [Oxfordshire]: Oxford University Press. ISBN 0-19-963820-9.
- ↑ Xie XS, Lu HP (June 1999). "Single-molecule enzymology". J. Biol. Chem. 274 (23): 15967–70. doi:10.1074/jbc.274.23.15967. PMID 10347141.
- ↑ Lu H (2004). "Single-molecule spectroscopy studies of conformational change dynamics in enzymatic reactions". Current pharmaceutical biotechnology 5 (3): 261–9. doi:10.2174/1389201043376887. PMID 15180547.
- ↑ Schnell J, Dyson H, Wright P (2004). "Structure, dynamics, and catalytic function of dihydrofolate reductase". Annual review of biophysics and biomolecular structure 33: 119–40. doi:10.1146/annurev.biophys.33.110502.133613. PMID 15139807.
- ↑ Gibson QH (1969). "Rapid mixing: Stopped flow". Methods Enzymol. Methods in Enzymology 16: 187–228. doi:10.1016/S0076-6879(69)16009-7. ISBN 978-0-12-181873-9.
- ↑ Duggleby RG (1995). "Analysis of enzyme progress curves by non-linear regression". Methods Enzymol. Methods in Enzymology 249: 61–90. doi:10.1016/0076-6879(95)49031-0. ISBN 978-0-12-182150-0. PMID 7791628.
- ↑ Murray JB, Dunham CM, Scott WG (January 2002). "A pH-dependent conformational change, rather than the chemical step, appears to be rate-limiting in the hammerhead ribozyme cleavage reaction". J. Mol. Biol. 315 (2): 121–30. doi:10.1006/jmbi.2001.5145. PMID 11779233.
- ↑ Michaelis L. and Menten M.L. Kinetik der Invertinwirkung Biochem. Z. 1913; 49:333–369 English translation Accessed 6 April 2007
- ↑ Stroppolo ME, Falconi M, Caccuri AM, Desideri A (2001). "Superefficient enzymes". Cell. Mol. Life Sci. 58 (10): 1451–60. doi:10.1007/PL00000788. PMID 11693526.
- ↑ Walsh R, Martin E, Darvesh S (Jan 2010). "A method to describe enzyme-catalyzed reactions by combining steady state and time course enzyme kinetic parameters". Biochim Biophys Acta 1800: 1–5. doi:10.1016/j.bbagen.2009.10.007.
- ↑ Beal, S. L. (1983). "Computation of the explicit solution to the Michaelis-Menten equation". Journal of Pharmacokinetics and Biopharmaceutics 11 (6): 641–657. doi:10.1007/BF01059062. PMID 6689584.
- ↑ Schnell, S.; Mendoza, C. (1997). "Closed Form Solution for Time-dependent Enzyme Kinetics". Journal of Theoretical Biology 187 (2): 207–212. doi:10.1006/jtbi.1997.0425.
- ↑ Goudar, CT; Sonnad, JR; Duggleby, RG (1999). "Parameter estimation using a direct solution of the integrated Michaelis-Menten equation" (PDF). Biochimica et Biophysica Acta 1429 (2): 377–83. doi:10.1016/s0167-4838(98)00247-7. PMID 9989222.
- ↑ Goudar, C. T.; Harris, S. K.; McInerney, M. J.; Suflita, J. M. (2004). "Progress curve analysis for enzyme and microbial kinetic reactions using explicit solutions based on the Lambert W function". Journal of Microbiological Methods 59 (3): 317–326. doi:10.1016/j.mimet.2004.06.013. PMID 15488275.
- ↑ Jones ME (1992). "Analysis of algebraic weighted least-squares estimators for enzyme parameters". Biochem. J. 288 (Pt 2): 533–8. PMC 1132043. PMID 1463456.
- ↑ Tseng SJ, Hsu JP (August 1990). "A comparison of the parameter estimating procedures for the Michaelis-Menten model". J. Theor. Biol. 145 (4): 457–64. doi:10.1016/S0022-5193(05)80481-3. PMID 2246896.
- ↑ Bravo IG, Busto F, De Arriaga D; et al. (September 2001). "A normalized plot as a novel and time-saving tool in complex enzyme kinetic analysis". Biochem. J. 358 (Pt 3): 573–83. PMC 1222113. PMID 11577687.
- ↑ Almaas E, Kovács B, Vicsek T, Oltvai ZN, Barabási AL (February 2004). "Global organization of metabolic fluxes in the bacterium Escherichia coli". Nature 427 (6977): 839–43. doi:10.1038/nature02289. PMID 14985762.
- ↑ Reed JL, Vo TD, Schilling CH, Palsson BO (2003). "An expanded genome-scale model of Escherichia coli K-12 (iJR904 GSM/GPR)". Genome Biol. 4 (9): R54. doi:10.1186/gb-2003-4-9-r54. PMC 193654. PMID 12952533.
- ↑ for a complete derivation, see here
- ↑ Dirr H, Reinemer P, Huber R (1994). "X-ray crystal structures of cytosolic glutathione S-transferases. Implications for protein architecture, substrate recognition and catalytic function". Eur. J. Biochem. 220 (3): 645–61. doi:10.1111/j.1432-1033.1994.tb18666.x. PMID 8143720.
- ↑ Stone SR, Morrison JF (July 1988). "Dihydrofolate reductase from Escherichia coli: the kinetic mechanism with NADPH and reduced acetylpyridine adenine dinucleotide phosphate as substrates". Biochemistry 27 (15): 5493–9. doi:10.1021/bi00415a016. PMID 3052577.
- ↑ Fisher PA (1994). "Enzymologic mechanism of replicative DNA polymerases in higher eukaryotes". Prog. Nucleic Acid Res. Mol. Biol. Progress in Nucleic Acid Research and Molecular Biology 47: 371–97. doi:10.1016/S0079-6603(08)60257-3. ISBN 978-0-12-540047-3. PMID 8016325.
- ↑ Akerman SE, Müller S (2003). "2-Cys peroxiredoxin PfTrx-Px1 is involved in the antioxidant defence of Plasmodium falciparum". Mol. Biochem. Parasitol. 130 (2): 75–81. doi:10.1016/S0166-6851(03)00161-0. PMID 12946843.
- ↑ Bravo IG, Barrallo S, Ferrero MA, Rodríguez-Aparicio LB, Martínez-Blanco H, Reglero A (September 2001). "Kinetic properties of the acylneuraminate cytidylyltransferase from Pasteurella haemolytica A2". Biochem. J. 358 (Pt 3): 585–98. PMC 1222114. PMID 11577688.
- ↑ Kraut J (1977). "Serine proteases: structure and mechanism of catalysis". Annu. Rev. Biochem. 46: 331–58. doi:10.1146/annurev.bi.46.070177.001555. PMID 332063.
- ↑ Ricard J, Cornish-Bowden A (July 1987). "Co-operative and allosteric enzymes: 20 years on". Eur. J. Biochem. 166 (2): 255–72. doi:10.1111/j.1432-1033.1987.tb13510.x. PMID 3301336.
- ↑ Ward WH, Fersht AR (July 1988). "Tyrosyl-tRNA synthetase acts as an asymmetric dimer in charging tRNA. A rationale for half-of-the-sites activity". Biochemistry 27 (15): 5525–30. doi:10.1021/bi00415a021. PMID 3179266.
- ↑ Helmstaedt K, Krappmann S, Braus GH (September 2001). "Allosteric Regulation of Catalytic Activity: Escherichia coli Aspartate Transcarbamoylase versus Yeast Chorismate Mutase". Microbiol. Mol. Biol. Rev. 65 (3): 404–21, table of contents. doi:10.1128/MMBR.65.3.404-421.2001. PMC 99034. PMID 11528003.
- ↑ Schirmer T, Evans PR (January 1990). "Structural basis of the allosteric behaviour of phosphofructokinase". Nature 343 (6254): 140–5. doi:10.1038/343140a0. PMID 2136935.
- ↑ Hill, A. V. The possible effects of the aggregation of the molecules of haemoglobin on its dissociation curves. J. Physiol. (Lond.), 1910 40, iv–vii.
- ↑ Hartley BS, Kilby BA (February 1954). "The reaction of p-nitrophenyl esters with chymotrypsin and insulin". Biochem. J. 56 (2): 288–97. PMC 1269615. PMID 13140189.
- 1 2 Fersht, Alan (1999). Structure and mechanism in protein science: a guide to enzyme catalysis and protein folding. San Francisco: W.H. Freeman. ISBN 0-7167-3268-8.
- ↑ Bender ML, Begué-Cantón ML, Blakeley RL; et al. (December 1966). "The determination of the concentration of hydrolytic enzyme solutions: alpha-chymotrypsin, trypsin, papain, elastase, subtilisin, and acetylcholinesterase". J. Am. Chem. Soc. 88 (24): 5890–913. doi:10.1021/ja00976a034. PMID 5980876.
- ↑ Cleland WW (January 2005). "The use of isotope effects to determine enzyme mechanisms". Arch. Biochem. Biophys. 433 (1): 2–12. doi:10.1016/j.abb.2004.08.027. PMID 15581561.
- ↑ Northrop D (1981). "The expression of isotope effects on enzyme-catalyzed reactions". Annu. Rev. Biochem. 50: 103–31. doi:10.1146/annurev.bi.50.070181.000535. PMID 7023356.
- ↑ Baillie T, Rettenmeier A (1986). "Drug biotransformation: mechanistic studies with stable isotopes". Journal of clinical pharmacology 26 (6): 448–51. doi:10.1002/j.1552-4604.1986.tb03556.x. PMID 3734135.
- ↑ Cleland WW (1982). "Use of isotope effects to elucidate enzyme mechanisms". CRC Crit. Rev. Biochem. 13 (4): 385–428. doi:10.3109/10409238209108715. PMID 6759038.
- ↑ Christianson DW, Cox JD (1999). "Catalysis by metal-activated hydroxide in zinc and manganese metalloenzymes". Annu. Rev. Biochem. 68: 33–57. doi:10.1146/annurev.biochem.68.1.33. PMID 10872443.
- ↑ Kraut D, Carroll K, Herschlag D (2003). "Challenges in enzyme mechanism and energetics". Annu. Rev. Biochem. 72: 517–71. doi:10.1146/annurev.biochem.72.121801.161617. PMID 12704087.
- ↑ Walsh, R.; Martin, E.; Darvesh, S. (2011). "Limitations of conventional inhibitor classifications". Integrative Biology (Royal Society of Chemistry) 3 (12): 1197–1201. doi:10.1039/c1ib00053e. PMID 22038120.
- ↑ Walsh, R.; Martin, E.; Darvesh, S. (2007). "A versatile equation to describe reversible enzyme inhibition and activation kinetics: Modeling β-galactosidase and butyrylcholinesterase". Biochimica et Biophysica Acta (BBA) - General Subjects 1770 (5): 733–746. doi:10.1016/j.bbagen.2007.01.001. PMID 17307293.
- ↑ Walsh, Ryan (2012). "Ch. 17. Alternative Perspectives of Enzyme Kinetic Modeling". In Ekinci, Deniz. Medicinal Chemistry and Drug Design (PDF). InTech. pp. 357–371. ISBN 978-953-51-0513-8.
- ↑ Koshland DE (February 1958). "Application of a Theory of Enzyme Specificity to Protein Synthesis". Proc. Natl. Acad. Sci. U.S.A. 44 (2): 98–104. doi:10.1073/pnas.44.2.98. PMC 335371. PMID 16590179.
- ↑ Hammes G (2002). "Multiple conformational changes in enzyme catalysis". Biochemistry 41 (26): 8221–8. doi:10.1021/bi0260839. PMID 12081470.
- ↑ Sutcliffe M, Scrutton N (2002). "A new conceptual framework for enzyme catalysis. Hydrogen tunnelling coupled to enzyme dynamics in flavoprotein and quinoprotein enzymes". Eur. J. Biochem. 269 (13): 3096–102. doi:10.1046/j.1432-1033.2002.03020.x. PMID 12084049.
- ↑ Henri V (1902). "Theorie generale de l'action de quelques diastases". Compt. Rend. Hebd. Acad. Sci. Paris 135: 916–9.
- ↑ Sørensen PL (1909). "Enzymstudien {II}. Über die Messung und Bedeutung der Wasserstoffionenkonzentration bei enzymatischen Prozessen" [Enzyme studies III: About the measurement and significance of the hydrogen ion concentration in enzymatic processes]. Biochem. Z. (in German) 21: 131–304.
- ↑ Michaelis L, Menten M (1913). "Die Kinetik der Invertinwirkung" [The Kinetics of Invertase Action]. Biochem. Z. (in German) 49: 333–369.; Michaelis L, Menten ML, Johnson KA, Goody RS (2011). "The original Michaelis constant: translation of the 1913 Michaelis-Menten paper". Biochemistry 50 (39): 8264–9. doi:10.1021/bi201284u. PMC 3381512. PMID 21888353.
- ↑ Briggs GE, Haldane JB (1925). "A Note on the Kinetics of Enzyme Action". The Biochemical Journal 19 (2): 339–339. PMC 1259181. PMID 16743508.
- ↑ Flomenbom O, Velonia K, Loos D, Masuo S, Cotlet M, Engelborghs Y, Hofkens J, Rowan AE, Nolte RJ, Van der Auweraer M, de Schryver FC, Klafter J (Feb 2005). "Stretched exponential decay and correlations in the catalytic activity of fluctuating single lipase molecules". Proceedings of the National Academy of Sciences of the United States of America 102 (7): 2368–2372. doi:10.1073/pnas.0409039102. PMC 548972. PMID 15695587.
- ↑ English BP, Min W, van Oijen AM, Lee KT, Luo G, Sun H, Cherayil BJ, Kou SC, Xie XS (Feb 2006). "Ever-fluctuating single enzyme molecules: Michaelis-Menten equation revisited". Nature Chemical Biology 2 (2): 87–94. doi:10.1038/nchembio759. PMID 16415859.
- ↑ Lu HP, Xun L, Xie XS (Dec 1998). "Single-molecule enzymatic dynamics". Science 282 (5395): 1877–1882. doi:10.1126/science.282.5395.1877. PMID 9836635.
- ↑ Xue X, Liu F, Ou-Yang ZC (Sep 2006). "Single molecule Michaelis-Menten equation beyond quasistatic disorder". Physical Review. E, Statistical, Nonlinear, and Soft Matter Physics 74 (3 Pt 1): 030902. doi:10.1103/PhysRevE.74.030902. PMID 17025584.
- ↑ Bevc S., Konc J., Stojan J., Hodošček M., Penca M., Matej Praprotnik M., Janežič D. (2011). "ENZO: A Web Tool for Derivation and Evaluation of Kinetic Models of Enzyme Catalyzed Reactions". PLoS ONE 6 (7): e22265. doi:10.1371/journal.pone.0022265. PMC 3139599. PMID 21818304. ENZO server
Further reading
Introductory
- Cornish-Bowden, Athel (2004). Fundamentals of enzyme kinetics (3rd ed.). London: Portland Press. ISBN 1-85578-158-1.
- Stevens, Lewis; Price, Nicholas C. (1999). Fundamentals of enzymology: the cell and molecular biology of catalytic proteins. Oxford [Oxfordshire]: Oxford University Press. ISBN 0-19-850229-X.
- Bugg, Tim (2004). Introduction to Enzyme and Coenzyme Chemistry. Cambridge, MA: Blackwell Publishers. ISBN 1-4051-1452-5.
Advanced
- Segel, Irwin H. (1993). Enzyme kinetics: behavior and analysis of rapid equilibrium and steady state enzyme systems (New ed.). New York: Wiley. ISBN 0-471-30309-7.
- Fersht, Alan (1999). Structure and mechanism in protein science: a guide to enzyme catalysis and protein folding. San Francisco: W.H. Freeman. ISBN 0-7167-3268-8.
- Santiago Schnell, Philip K. Maini (2004). "A century of enzyme kinetics: Reliability of the KM and vmax estimates" (PDF). Comments on Theoretical Biology 8 (2–3): 169–87. doi:10.1080/08948550302453.
- Walsh, Christopher (1979). Enzymatic reaction mechanisms. San Francisco: W. H. Freeman. ISBN 0-7167-0070-0.
- Cleland, William Wallace; Cook, Paul (2007). Enzyme kinetics and mechanism. New York: Garland Science. ISBN 0-8153-4140-7.
External links
- Animation of an enzyme assay — Shows effects of manipulating assay conditions
- MACiE — A database of enzyme reaction mechanisms
- ENZYME — Expasy enzyme nomenclature database
- ENZO — Web application for easy construction and quick testing of kinetic models of enzyme catalyzed reactions.
- ExCatDB — A database of enzyme catalytic mechanisms
- BRENDA — Comprehensive enzyme database, giving substrates, inhibitors and reaction diagrams
- SABIO-RK — A database of reaction kinetics
- Joseph Kraut's Research Group, University of California San Diego — Animations of several enzyme reaction mechanisms
- Symbolism and Terminology in Enzyme Kinetics — A comprehensive explanation of concepts and terminology in enzyme kinetics
- An introduction to enzyme kinetics — An accessible set of on-line tutorials on enzyme kinetics
- Enzyme kinetics animated tutorial — An animated tutorial with audio
| ||||||||||||||||||