Eicosanoid

In biochemistry, eicosanoids (preferred IUPAC name icosanoids) are signaling molecules made by oxidation of 20-carbon fatty acids. They exert complex control over many bodily systems; mainly in growth during and after physical activity, inflammation or immunity after the intake of toxic compounds and pathogens, and as messengers in the central nervous system. Many are classified as hormones. The networks of controls that depend upon eicosanoids are among the most complex in the human body.
Eicosanoids are derived from either omega-3 (ω-3) or omega-6 (ω-6) fatty acids. In general, the ω-6 eicosanoids are pro-inflammatory; ω-3s are much less so. The amounts and balance of these fats in a person's diet will affect the body's eicosanoid-controlled functions, with effects on cardiovascular disease, triglycerides, blood pressure, and arthritis.
There are multiple subfamilies of eicosanoids, including the prostaglandins, thromboxanes, and leukotrienes, as well as the lipoxins and eoxins, and others. For each, there are two or three separate series, derived from either an ω-3 or an ω-6 EFA. These series' different activities largely explain the health effects of ω-3 and ω-6 fats.[1][2][3][4]
Nomenclature
- See related detail at Essential Fatty Acid Interactions—Nomenclature
"Eicosanoid" (eicosa-, Greek for "twenty"; see icosahedron) is the collective term[5] for oxygenated derivatives of three different 20-carbon fatty acids:
- Eicosapentaenoic acid (EPA), an ω-3 fatty acid with 5 double bonds;
- Arachidonic acid (AA), an ω-6 fatty acid, with 4 double bonds;
- Dihomo-gamma-linolenic acid (DGLA), an ω-6, with 3 double bonds.
Current usage limits the term eicosanoid to leukotrienes (LT), eoxins (EX), and three types of prostanoids—prostaglandins (PG), prostacyclins (PGI) and thromboxanes (TX). This is the definition used in this article. However, several other classes can technically be termed eicosanoid, including the hepoxilins, resolvins, isofurans, isoprostanes, lipoxins, epi-lipoxins, epoxyeicosatrienoic acids (EETs) and endocannabinoids. LTs and prostanoids are sometimes termed 'classic eicosanoids'[6][7][8] in contrast to the 'novel', 'eicosanoid-like' or 'nonclassic eicosanoids'.[9][10][11][12]
A particular eicosanoid is denoted by a four-character abbreviation, composed of:
- Its two-letter abbreviation (above),[13]
- One A-B-C sequence-letter;[14] and
- A subscript, indicating the number of double bonds.
Examples are:
- The EPA-derived prostanoids have three double bonds, (e.g., PGG3, PGH3, PGI3, TXA3) while its leukotrienes have five, (LTB5).
- The AA-derived prostanoids have two double bonds, (e.g., PGG2, PGH2, PGI2, TXA2) while its leukotrienes have four, (LTB4).
Furthermore, stereochemistry may differ among the pathways, indicated by Greek letters, e.g. for (PGF2α).
Biosynthesis
Three families of enzymes catalyze fatty acid oxygenation to produce the eicosanoids:
- Cyclooxygenase, or COX, generates the prostanoids from arachidonic acid.
- Lipoxygenase, or LOX, in several forms. 5-Lipoxygenase (5-LO) generates the leukotrienes from arachidonic acid and via transcellular biosynthesis is also involved in lipoxin generation. 15-Lipoxygenase generates the eoxins.
- Epoxygenases are a large number of cytochrome P450 enzymes which generate nonclassic eicosanoid epoxides viz., the epoxyeicosatrienoic acids derived from arachidonic acid and the epoxyeicoatetraenoic acids derived from eicosapentaenoic acid.
Eicosanoids are not stored within cells, but are synthesized as required. They derive from the fatty acids that make up the cell membrane and nuclear membrane.
Eicosanoid biosynthesis begins when a cell is activated by mechanical trauma, cytokines, growth factors or other stimuli. (The stimulus may even be an eicosanoid from a neighboring cell; the pathways are complex.) This triggers the release of a phospholipase at the cell membrane. The phospholipase travels to the nuclear membrane. There, the phospholipase catalyzes ester hydrolysis of phospholipid (by phospholipase A2) or diacylglycerol (by phospholipase C). This frees a 20-carbon fatty acid. This hydrolysis appears to be the rate-determining step for eicosanoid formation.
The fatty acids may be released by any of several phospholipases. Of these, type IV cytosolic phospholipase A2 (cPLA2) is the key actor, as cells lacking cPLA2 are, in general, devoid of eicosanoid synthesis. The phospholipase cPLA2 is specific for phospholipids that contain AA, EPA or GPLA at the SN2 position. Interestingly, cPLA2 may also release the lysophospholipid that becomes platelet-activating factor.[15]
Peroxidation and reactive oxygen species
Next, the free fatty acid is oxygenated along any of several pathways; see the Pathways table. The eicosanoid pathways (via lipoxygenase or COX) add molecular oxygen (O2). Although the fatty acid is symmetric, the resulting eicosanoids are chiral; the oxidations proceed with high stereoselectivity (enzymatic oxidations are considered practically stereospecific).
The oxidation of lipids is hazardous to cells, particularly when close to the nucleus. There are elaborate mechanisms to prevent unwanted oxidation. COX, the lipoxygenases and the phospholipases are tightly controlled—there are at least eight proteins activated to coordinate generation of leukotrienes. Several of these exist in multiple isoforms.[4]
Oxidation by either COX or lipoxygenase releases reactive oxygen species (ROS) and the initial products in eicosanoid generation are themselves highly reactive peroxides. LTA4 can form adducts with tissue DNA. Other reactions of lipoxygenases generate cellular damage; murine models implicate 15-lipoxygenase in the pathogenesis of atherosclerosis.[16][17] The oxidation in eicosanoid generation is compartmentalized; this limits the peroxides' damage. The enzymes that are biosynthetic for eicosanoids (e.g., glutathione-S-transferases, epoxide hydrolases, and carrier proteins) belong to families whose functions are involved largely with cellular detoxification. This suggests that eicosanoid signaling might have evolved from the detoxification of ROS.
The cell must realize some benefit from generating lipid hydroperoxides close-by its nucleus. PGs and LTs may signal or regulate DNA-transcription there; LTB4 is ligand for PPARα.[2] (See diagram at PPAR).
 |
 |
 | |
| Prostaglandin E1. The 5-member ring is characteristic of the class. | Thromboxane A2. Oxygens have moved into the ring. |
Leukotriene B4. Note the 3 conjugated double bonds. | |
 |
 | ||
| Prostacyclin I2. The second ring distinguishes it from the prostaglandins. | Leukotriene E4, an example of a cysteinyl leukotriene. | ||
Prostanoid pathways
Cyclooxygenase (COX) catalyzes the conversion of the free fatty acids to prostanoids by a two-step process. First, two molecules of O2 are added as two peroxide linkages, and a 5-member carbon ring is forged near the middle of the fatty acid chain. This forms the short-lived, unstable intermediate Prostaglandin G (PGG). Next, one of the peroxide linkages sheds a single oxygen, forming PGH. (See diagrams and more detail of these steps at Cyclooxygenase).
All three classes of prostanoids originate from PGH. All have distinctive rings in the center of the molecule. They differ in their structures. The PGH compounds (parents to all the rest) have a 5-carbon ring, bridged by two oxygens (a peroxide.) As the example in Structures of Selected Eicosanoids figure shows, the derived prostaglandins contain a single, unsaturated 5-carbon ring. In prostacyclins, this ring is conjoined to another oxygen-containing ring. In thromboxanes the ring becomes a 6-member ring with one oxygen. The leukotrienes do not have rings. (See more detail, including the enzymes involved, in diagrams at Prostanoid.)
Several drugs lower inflammation by blocking prostanoid synthesis; see detail at Cyclooxygenase, Aspirin and NSAID.
Hydroxyeicosatetraenoate (HETE) and leukotriene (LT) pathways
See Leukotriene#Biosynthesis, Hydroxyeicosatetraenoic acid, and Eoxin#Human biosynthesis.
The enzyme 5-lipoxygenase (5-LO or ALOX5) uses 5-lipoxygenase activating protein (FLAP) to convert arachidonic acid into 5-hydroperoxyeicosatetraenoic acid (5-HPETE), which if not further metabolized by the enzyme LTA synthase, is rapidly reduces to 5-hydroxyeicosatetraenoic acid (5-HETE) by ubiquitous cellular glutathione-dependent peroxidases.[18] The enzyme LTA synthase acts on 5-HPETE to convert it into leukotriene A4 (LTA4), which may be converted into LTB4 by the enzyme leukotriene A4 epoxide hydrolase. Eosinophils, mast cells, and alveolar macrophages use the enzyme leukotriene C4 synthase to conjugate glutathione with LTA4 to make LTC4, which is transported outside the cell, where a glutamic acid moiety is removed from it to make LTD4. The leukotriene LTD4 is then cleaved by dipeptidases to make LTE4. The leukotrienes LTC4, LTD4 and LTE4 all contain cysteine and are collectively known as the cysteinyl leukotrienes.
The enzyme arachidonate 12-lipoxygenase (12-LO or ALOX12) metabolizes arachidonic acid to the S stereoisomer of 12-hydroperoxyeicosatetraenoic acid (5-HPETE) which is rapidly reduced by cellular peroxidases to the S stereoisomer of 12-hydroxyeicosatetraenoic acid (12-HETE) or further metabolized to hepoxilins (Hx) such as HxA3 and HxB.[19][20]
The enzymes 15-lipoxygenase-1 (15-LO-1 or ALOX15) and 15-lipoxygenase-2 (15-LO-2, ALOX15B) metabolize arachidonic acid to the S stereoisomer of 15-Hydroperoxyeicosatetraenoic acid (15-HPETE) which is rapidly reduced by cellular peroxidases to the S stereoisomer of 15-Hydroxyicosatetraenoic acid (15-HETE).[21][22]
A subset of Cytochrome P450 (CYP450) microsome-bound ω-hydroxylases (see 20-Hydroxyeicosatetraenoic acid) metabolize arachidonic acid to 20-Hydroxyeicosatetraenoic acid (20-HETE) and 19-hydroxyeicosatetraenoic acid by an omega oxidation reaction.[23]
Epoxyeicosanoid pathway
The human cytochrome P450 (CYP) epoxygenases, CYP1A1, CYP1A2, CYP2C8, CYP2C9, CYP2C18, CYP2C19, CYP2E1, CYP2J2, and CYP2S1 metabolize arachidonic acid to the non-classic Epoxyeicosatrienoic acids (EETs) by coverting one of the fatty acid's double bonds to its epoxide to form one or more of the following EETs, 14,15-ETE, 11,12-EET, 8,9-ETE, and 4,5-ETE.[24][25] 14,15-EET and 11,12-EET are the major EETs produced by mammalian, including human, tissues.[25][25][26][27][28][29] The same CYPs but also CYP4A1, CYP4F8, and CYP4F12 metabolize eicosapentaenoic acid to five epoxides viz., 17,18-, 14,15-, 11,12-. 8,9-, and 5,6-epoxy-eicsatetraenoic acids.[30]
Function and pharmacology
| PGD2 | Promotion of sleep | TXA2 | Stimulation of platelet aggregation; vasoconstriction |
| PGE2 | Smooth muscle contraction; inducing pain, heat, fever; bronchoconstriction |
15d-PGJ2 | Adipocyte differentiation |
| PGF2α | Uterine contraction | LTB4 | Leukocyte chemotaxis |
| PGI2 | Inhibition of platelet aggregation; vasodilation; embryo implantation |
Cysteinyl-LTs | Anaphylaxis; bronchial smooth muscle contraction. |
| †Shown eicosanoids are AA-derived; in general, EPA-derived have weaker activity | |||
Eicosanoids exert complex control over many bodily systems, mainly in inflammation or immunity, and as messengers in the central nervous system. They are found in most living things. In humans, eicosanoids are local hormones that are released by most cells, act on that same cell or nearby cells (i.e., they are autocrine and paracrine mediators), and then are rapidly inactivated.
Eicosanoids have a short half-life, ranging from seconds to minutes. Dietary antioxidants inhibit the generation of some inflammatory eicosanoids, e.g. trans-resveratrol against thromboxane and some leukotrienes.[31] Most eicosanoid receptors are members of the G protein-coupled receptor superfamily; see the Receptors table or the article eicosanoid receptors.
Leukotrienes:
|
Prostanoids:
|
The ω-3 and ω-6 series
| “ | The reduction in AA-derived eicosanoids and the diminished activity of the alternative products generated from ω-3 fatty acids serve as the foundation for explaining some of the beneficial effects of greater ω-3 intake. | ” | |
| — Kevin Fritsche, Fatty Acids as Modulators of the Immune Response[32] | |||
Arachidonic acid (AA; 20:4 ω-6) sits at the head of the 'arachidonic acid cascade'—more than twenty different eicosanoid-mediated signaling paths controlling a wide array of cellular functions, especially those regulating inflammation, immunity and the central nervous system.[3]
In the inflammatory response, two other groups of dietary fatty acids form cascades that parallel and compete with the arachidonic acid cascade. EPA (20:5 ω-3) provides the most important competing cascade. DGLA (20:3 ω-6) provides a third, less prominent cascade. These two parallel cascades soften the inflammatory effects of AA and its products. Low dietary intake of these less-inflammatory fatty acids, especially the ω-3s, has been linked to several inflammation-related diseases, and perhaps some mental illnesses.
The U.S. National Institutes of Health and the National Library of Medicine state that there is 'A' level evidence that increased dietary ω-3 improves outcomes in hypertriglyceridemia, secondary cardiovascular disease prevention and hypertension. There is 'B' level evidence ('good scientific evidence') for increased dietary ω-3 in primary prevention of cardiovascular disease, rheumatoid arthritis and protection from ciclosporin toxicity in organ transplant patients. They also note more preliminary evidence showing that dietary ω-3 can ease symptoms in several psychiatric disorders.[33]
Besides the influence on eicosanoids, dietary polyunsaturated fats modulate immune response through three other molecular mechanisms. They
(a) alter membrane composition and function, including the composition of lipid rafts;
(b) change cytokine biosynthesis and (c) directly activate gene transcription.[32] Of these, the action on eicosanoids is the best explored.
Mechanisms of ω-3 action
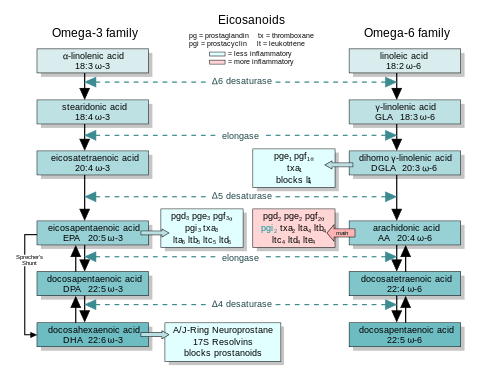
In general, the eicosanoids derived from AA promote inflammation, and those from EPA and from GLA (via DGLA) are less inflammatory, or inactive, or even anti-inflammatory and pro-resolving.
The figure shows the ω-3 and -6 synthesis chains, along with the major eicosanoids from AA, EPA and DGLA.
Dietary ω-3 and GLA counter the inflammatory effects of AA's eicosanoids in three ways, along the eicosanoid pathways:
- Displacement—Dietary ω-3 decreases tissue concentrations of AA, so there is less to form ω-6 eicosanoids.
- Competitive inhibition—DGLA and EPA compete with AA for access to the cyclooxygenase and lipoxygenase enzymes. So the presence of DGLA and EPA in tissues lowers the output of AA's eicosanoids.
- Counteraction—Some DGLA and EPA derived eicosanoids counteract their AA derived counterparts.
Role in inflammation
Since antiquity, the cardinal signs of inflammation have been known as: calor (warmth), dolor (pain), tumor (swelling) and rubor (redness). The eicosanoids are involved with each of these signs.
Redness—An insect's sting will trigger the classic inflammatory response. Short acting vasoconstrictors — TXA2—are released quickly after the injury. The site may momentarily turn pale. Then TXA2 mediates the release of the vasodilators PGE2 and LTB4. The blood vessels engorge and the injury reddens.
Swelling—LTB4 makes the blood vessels more permeable. Plasma leaks out into the connective tissues, and they swell. The process also loses pro-inflammatory cytokines.
Pain—The cytokines increase COX-2 activity. This elevates levels of PGE2, sensitizing pain neurons.
Heat—PGE2 is also a potent pyretic agent. Aspirin and NSAIDS—drugs that block the COX pathways and stop prostanoid synthesis—limit fever or the heat of localized inflammation.
| Medicine | Type | Medical condition or use |
|---|---|---|
| Alprostadil | PGE1 | Erectile dysfunction, maintaining a patent ductus arteriosus in the fetus |
| Beraprost | PGI1 analog | Pulmonary hypertension, avoiding reperfusion injury |
| Bimatoprost | PGF2α analog | Glaucoma, ocular hypertension |
| Carboprost | PGF2α analog | Labor induction, abortifacient in early pregnancy |
| Dinoprostone | PGE2 | Labor induction |
| Iloprost | PGI2 analog | Pulmonary arterial hypertension |
| Latanoprost | PGF2α analog | Glaucoma, ocular hypertension |
| Misoprostol | PGE1 analog | Stomach ulcers, labor induction, abortifacient |
| Montelukast | LT receptor antagonist |
Asthma, seasonal allergies |
| Travoprost | PGF2α analog | Glaucoma, ocular hypertension |
| Treprostinil | PGI analog | Pulmonary hypertension |
| U46619 | Longer lived TX analog |
Research only |
| Zafirlukast | LT receptor antagonist |
Asthma |
Action of prostanoids
- Main articles: Prostaglandin, Prostacyclin and Thromboxane
Prostanoids mediate local symptoms of inflammation: vasoconstriction or vasodilation, coagulation, pain and fever. Inhibition of cyclooxygenase, specifically the inducible COX-2 isoform, is the hallmark of NSAIDs (non-steroidal anti-inflammatory drugs), such as aspirin. COX-2 is responsible for pain and inflammation, while COX-1 is responsible for platelet clotting actions.
Prostanoids activate the PPARγ members of the steroid/thyroid family of nuclear hormone receptors, directly influencing gene transcription.[34]
Action of leukotrienes
Leukotrienes play an important role in inflammation. There is a neuroendocrine role for LTC4 in luteinizing hormone secretion.[35] LTB4 causes adhesion and chemotaxis of leukocytes and stimulates aggregation, enzyme release, and generation of superoxide in neutrophils.[36] Blocking leukotriene receptors can play a role in the management of inflammatory diseases such as asthma (by the drugs montelukast and zafirlukast), psoriasis, and rheumatoid arthritis.
The slow reacting substance of anaphylaxis comprises the cysteinyl leukotrienes. These have a clear role in pathophysiological conditions such as asthma, allergic rhinitis and other nasal allergies, and have been implicated in atherosclerosis and inflammatory gastrointestinal diseases.[37] They are potent bronchoconstrictors, increase vascular permeability in postcapillary venules, and stimulate mucus secretion. They are released from the lung tissue of asthmatic subjects exposed to specific allergens and play a pathophysiological role in immediate hypersensitivity reactions.[36] Along with PGD, they function in effector cell trafficking, antigen presentation, immune cell activation, matrix deposition, and fibrosis.[38]
Action of epoxyeicosanoids
The Epoxy eicostrienoic acids or EETs and, it is in general presumed if not clearly shown, the epoxy eicosatetraenoic acids have vasodilating actions on heart, kidney and other blood vessels as well as on the kidney's reabsorption of sodium and water that act to reduce blood pressure and ischemic and other injuries to the heart, brain, and other tissues; they may also act to reduce inflammation, promote the growth and metastasis of certain tumors, promote the growth of new blood vessels, in the central nervous system regulate the release of neuropeptide hormones, and in the peripheral nervous system inhibit or reduce pain perception.[24][25][27]
History
In 1930, gynecologist Raphael Kurzrok and pharmacologist Charles Leib characterized prostaglandin as a component of semen. Between 1929 and 1932, Burr and Burr showed that restricting fat from animal's diets led to a deficiency disease, and first described the essential fatty acids.[39] In 1935, von Euler identified prostaglandin. In 1964, Bergström and Samuelsson linked these observations when they showed that the "classical" eicosanoids were derived from arachidonic acid, which had earlier been considered to be one of the essential fatty acids.[40] In 1971, Vane showed that aspirin and similar drugs inhibit prostaglandin synthesis.[41] Von Euler received the Nobel Prize in medicine in 1970, which Samuelsson, Vane, and Bergström also received in 1982. E. J. Corey received it in chemistry in 1990 largely for his synthesis of prostaglandins.
References
- ↑ DeCaterina, R and Basta, G (June 2001). "n-3 Fatty acids and the inflammatory response – biological background" (PDF). European Heart Journal Supplements. 3, Suppl D: D42–D49. doi:10.1016/S1520-765X(01)90118-X. Retrieved 2006-02-10.
- 1 2 Funk, Colin D. (30 November 2001). "Prostaglandins and Leukotrienes: Advances in Eicosanoid Biology". Science 294 (5548): 1871–1875. doi:10.1126/science.294.5548.1871. PMID 11729303. Retrieved 2007-01-08.
- 1 2 Piomelli, Daniele (2000). "Arachidonic Acid". Neuropsychopharmacology: The Fifth Generation of Progress. Retrieved 2006-03-03.
- 1 2 Soberman, Roy J. and Christmas, Peter (2003). "The organization and consequences of eicosanoid signaling". J. Clin. Invest 111 (8): 1107–1113. doi:10.1172/JCI200318338. PMC 152944. PMID 12697726. Retrieved 2007-01-05.
- ↑ Beare-Rogers (2001). "IUPAC Lexicon of Lipid Nutrition" (PDF). Retrieved June 1, 2006.
- ↑ Van Dyke TE, Serhan CN (2003). "Resolution of inflammation: a new paradigm for the pathogenesis of periodontal diseases". J. Dent. Res. 82 (2): 82–90. doi:10.1177/154405910308200202. PMID 12562878.
- ↑ Serhan CN, Gotlinger K, Hong S, Arita M (2004). "Resolvins, docosatrienes, and neuroprotectins, novel omega-3-derived mediators, and their aspirin-triggered endogenous epimers: an overview of their protective roles in catabasis". Prostaglandins Other Lipid Mediat. 73 (3–4): 155–72. doi:10.1016/j.prostaglandins.2004.03.005. PMID 15290791.
- ↑ Anderle P, Farmer P, Berger A, Roberts MA (2004). "Nutrigenomic approach to understanding the mechanisms by which dietary long-chain fatty acids induce gene signals and control mechanisms involved in carcinogenesis". Nutrition (Burbank, Los Angeles County, Calif.) 20 (1): 103–8. doi:10.1016/j.nut.2003.09.018. PMID 14698023.
- ↑ Evans AR, Junger H, Southall MD; et al. (2000). "Isoprostanes, novel eicosanoids that produce nociception and sensitize rat sensory neurons". J. Pharmacol. Exp. Ther. 293 (3): 912–20. PMID 10869392.
- ↑ O'Brien WF, Krammer J, O'Leary TD, Mastrogiannis DS (1993). "The effect of acetaminophen on prostacyclin production in pregnant women". Am. J. Obstet. Gynecol. 168 (4): 1164–9. doi:10.1016/0002-9378(93)90362-m. PMID 8475962.
- ↑ Behrendt H, Kasche A, Ebner von Eschenbach C, Risse U, Huss-Marp J, Ring J (2001). "Secretion of proinflammatory eicosanoid-like substances precedes allergen release from pollen grains in the initiation of allergic sensitization". Int. Arch. Allergy Immunol. 124 (1–3): 121–5. doi:10.1159/000053688. PMID 11306946.
- ↑ Sarau HM, Foley JJ, Schmidt DB; et al. (1999). "In vitro and in vivo pharmacological characterization of SB 201993, an eicosanoid-like LTB4 receptor antagonist with anti-inflammatory activity". Prostaglandins Leukot. Essent. Fatty Acids 61 (1): 55–64. doi:10.1054/plef.1999.0074. PMID 10477044.
- ↑ Prostacyclin—PGI—was previously classified as prostaglandin and retains its old identifier.
- ↑ Eicosanoids with different letters have placement of double-bonds and different functional groups attached to the molecular skeleton. Letters indicate roughly the order the eicosanoids were first described in the literature. For diagrams for PG [A–H] see Cyberlipid Center. "Prostanoids". Retrieved 2007-02-05.
- 1 2 University of Kansas Medical Center (2004). "Eicosanoids and Inflammation" (PDF). Retrieved 2007-01-05.
- ↑ Cyrus, Tillmann; Witztum, Joseph L.; Rader, Daniel J.; Tangirala, Rajendra; Fazio, Sergio; Linton, Macrae F.; Funk, Colin D. (June 1999). "Disruption of the 12/15-lipoxygenase gene diminishes atherosclerosis in apo E–deficient mice". J Clin Invest 103 (11): 1597–1604n. doi:10.1172/JCI5897. PMC 408369. PMID 10359569.
- ↑ Schewe T. (Mar–Apr 2002). "15-lipoxygenase-1: a prooxidant enzyme". Biol Chem. 383 (3–4): 365–74. doi:10.1515/BC.2002.041. PMID 12033428.
- ↑ Prog Lipid Res. 2013 Oct;52(4):651-65. doi: 10.1016/j.plipres.2013.09.001. Epub 2013 Sep 19. Review.PMID 24056189
- ↑ Br J Pharmacol. 2009 Oct;158(4):972-81. doi:10.1111/j.1476-5381.2009.00168.x Epub 2009 Apr 3>
- ↑ Prog Lipid Res. 2011 January; 50(1): 115–131. Published online 2010 October 21. doi: 10.1016/j.plipres.2010.10.005P MCID PMC3012140
- ↑ Gene. 2015 Nov 15;573(1):1-32. doi: 10.1016/j.gene.2015.07.073 Epub 2015 Jul 26. Review.PMID 26216303
- ↑ Prostaglandins Leukot Essent Fatty Acids. 2007 Aug;77(2):67-77. Epub 2007 Sep 14. Review. PMID 17869078
- ↑ Kroetz DL, Xu F (2005). "Regulation and inhibition of arachidonic acid omega-hydroxylases and 20-HETE formation". Annual Review of Pharmacology and Toxicology 45: 413–38. doi:10.1146/annurev.pharmtox.45.120403.100045. PMID 15822183.
- 1 2 Br J Clin Pharmacol. 2015 Jul;80(1):28-44. doi: 10.1111/bcp.12603. Epub 2015 Jun 1.PMID 25655310
- 1 2 3 4 Biochim Biophys Acta 2015 Apr;1851(4):356-65. doi: 10.1016/j.bbalip.2014.07.020. Epub 2014 Aug 2. Review.PMID 25093613
- ↑ Arch Biochem Biophys. 2008 Mar 15;471(2):116-25. doi: 10.1016/j.abb.2008.01.002. Epub 2008 Jan 11.PMID 18206980
- 1 2 Pharmacol Ther. 2014 Nov;144(2):134-61. doi: 10.1016/j.pharmthera.2014.05.011. Epub 2014 Jun 2. Review.PMID 24882266
- ↑ Basic Res Cardiol. 2013 Jan;108(1):319. doi: 10.1007/s00395-012-0319-8. Epub 2012 Dec 7. PMID 23224081
- ↑ Pharmacol Rev. 2014 Oct;66(4):1106-40. doi: 10.1124/pr.113.007781. Review.PMID 25244930
- ↑ Prostaglandins Other Lipid Mediat. 2011 Nov;96(1-4):99-108. doi: 10.1016/j.prostaglandins.2011.09.001. Epub 2011 Sep 16. Review.PMID 21945326
- ↑ Pace-Asciak CR, Hahn S, Diamandis EP, Soleas G, Goldberg DM. (31 March 1995). "The red wine phenolics trans-resveratrol and quercetin block human platelet aggregation and eicosanoid synthesis: implications for protection against coronary heart disease". Clin Chim Acta 235 (2): 207–19. doi:10.1016/0009-8981(95)06045-1. PMID 7554275.
- 1 2 Fritsche, Kevin (August 2006). "Fatty Acids as Modulators of the Immune Response". Annual Review of Nutrition 26: 45–73. doi:10.1146/annurev.nutr.25.050304.092610. PMID 16848700. Retrieved 2007-01-11.
- ↑ National Institute of Health (2005-08-01). "Omega-3 fatty acids, fish oil, alpha-linolenic acid". Archived from the original on May 3, 2006. Retrieved March 26, 2006.
- ↑ Bos C, Richel D, Ritsema T, Peppelenbosch M, Versteeg H (2004). "Prostanoids and prostanoid receptors in signal transduction". Int J Biochem Cell Biol 36 (7): 1187–205. doi:10.1016/j.biocel.2003.08.006. PMID 15109566.
- ↑ Samuelsson, SE Dahlen, JA Lindgren, CA Rouzer, and CN Serhan (1987). "Leukotrienes and lipoxins: structures, biosynthesis, and biological effects". Science 237 (4819): 1171–1176. doi:10.1126/science.2820055. PMID 2820055. Retrieved 2007-01-22.
- 1 2 Samuelsson B (May 1983). "Leukotrienes: mediators of immediate hypersensitivity reactions and inflammation". Science 220 (4597): 568–575. doi:10.1126/science.6301011. PMID 6301011.
- ↑ Capra V (2004). "Molecular and functional aspects of human cysteinyl leukotriene receptors". Pharmacol Res 50 (1): 1–11. doi:10.1016/j.phrs.2003.12.012. PMID 15082024.
- ↑ Boyce J (2005). "Eicosanoid mediators of mast cells: receptors, regulation of synthesis, and pathobiologic implications". Chem Immunol Allergy. Chemical Immunology and Allergy 87: 59–79. doi:10.1159/000087571. ISBN 3-8055-7948-9. PMID 16107763.
- ↑ Burr, G.O. and Burr, M.M. (1930). "On the nature and role of the fatty acids essential in nutrition" (PDF). J. Biol. Chem. 86 (587). Retrieved 2007-01-17.
- ↑ Bergström, S., Danielsson, H. and Samuelsson, B. (1964). "The enzymatic formation of prostaglandin E2 from arachidonic acid". Biochim. Biophys. Acta 90 (207): 207–10. doi:10.1016/0304-4165(64)90145-x. PMID 14201168.
- ↑ Vane, J. R. (June 23, 1971). "Inhibition of prostaglandin synthesis as a mechanism of action for aspirin-like drugs". Nature New Biol. 231 (25): 232–5. doi:10.1038/newbio231232a0. PMID 5284360.
External links
- Eicosanoids at the US National Library of Medicine Medical Subject Headings (MeSH)
| ||||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||