Dire wolf
| Dire wolf Temporal range: Rancholabrean, 0.25–0.010 Ma | |
|---|---|
 | |
| Mounted skeleton | |
| Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Chordata |
| Class: | Mammalia |
| Order: | Carnivora |
| Family: | Canidae |
| Genus: | Canis |
| Species: | †C. dirus |
| Binomial name | |
| †Canis dirus Leidy, 1858.[1] | |
| Synonyms | |
The dire wolf (Canis dirus, "fearsome dog") is an extinct carnivorous mammal of the genus Canis, roughly the size of the extant gray wolf, but with a heavier build. It evolved in the New World and was its most derived genus Canis species.[8] Canis dirus is assigned to the Rancholabrean land mammal age of North America (240,000–10,000 years BP) and was among the many large carnivores and megaherbivores that became extinct in North and South America near the end of the Pleistocene epoch. Its ending is associated with the Quaternary extinction event.
Taxonomy

Canis dirus[10] was named by Joseph Leidy in 1858.[1] In 1984, Kurten recognized a geographic variation within the North American dire wolf populations and proposed two subspecies: Canis dirus guildayi for specimens from California and Mexico with shorter limbs and longer teeth, and Canis dirus dirus for specimens east of the North American Continental Divide with longer limbs and shorter teeth.[11][12][13]
The first type specimen was originally found in the summer of 1854 at Evansville, Indiana when the Ohio River was quite low. The specimen, a fossilized jawbone, was obtained by Joseph Granville Norwood from an Evansville collector named Francis A. Linck. Leidy determined the specimen represented an extinct species of wolf and reported it under the name of Canis primaevus.[2] Norwood's letters to Leidy are preserved along with the type specimen at the Academy of Natural Sciences. Canis primaevus (Leidy 1854) was later renamed Canis indianensis (Leidy 1869) when Leidy discovered that the name Canis primaevus had previously been used overseas.[3]
Canis indianensis (Leidy 1869) was first associated with Canis dirus (Leidy 1858) by Allen in 1876 along with his discovery of Canis mississippiensis. As there were so few pieces of these 3 specimens, it was thought best to leave each specimen listed under these 3 provisional names until more material could be provided to show their relationship.[4] Canis indianensis (Leidy 1869) was declared a subjective synonym with Canis dirus according to a taxonomic opinion of Troxell in 1915.[14] In 1918, Merriam studied these fossils and proposed consolidating their names under the separate genus Aenocyon (from Aenos:terrible and cyon:wolf) to become Aenocyon dirus,[6] however not everyone agreed with moving away from genus Canis.[15] Canis ayersi (Sellards 1916) and Aenocyon dirus (Merriam 1918) were subjective synonyms with Canis dirus according to a taxonomic opinion of Lundelius in 1972.[16][17] All of the named taxa were subjective synonyms with Canis dirus according to a taxonomic opinion by Nowak in 1979,[18] except for Aenocyon dirus nebrascensis.
Evolution

The fossil record suggests that Feliforms and caniforms emerged within the super-family Carnivoramorpha 43 million years before present (YBP).[20] The caniforms included the fox-like Leptocyon genus whose various species existed from 34 million YBP before branching 11.9 million YBP into vulpes (foxes) and canini (canines). The Eucyon genus diverged 6.2 million YBP towards Canis ferox, which diverged 5 million YBP towards Canis lepophagus, which diverged 3.5 million YBP towards the wolf-like canids.[19]:174–5
North America
The sudden appearance in North America during the early Pleistocene of the large Canis armbrusteri (Armbruster's wolf) suggests that this was an immigrant from Asia, as was Canis lupus (Gray wolf) later in the Pleistocene.[19]:144 In China, the Pliocene wolf Canis chihliensis was a sister taxon of the lupus clade and may have been the ancestor for both Canis armbrusteri and Canis lupus.[19]:146
The late Irvingtonian wolf Aenocyon dirus nebrascensis (Frick 1930 undescribed) from the Hay Springs area in Sheridan County, Nebraska, may represent the earliest record of the largely Rancholabrean Canis dirus, according to both Frick[7] and Tedford.[19]:146 Nowak referred to the material as Canis armbrusteri,[18]:93 however Tedford described the material and noted that although these exhibited some morphological characteristics of both, he referred to the material as Canis dirus.[19]:146 There is good evidence of the evolution of Canis dirus from Canis armbrusteri. These taxa share a number of characteristics (synapomorphy), which suggests an origin of Canis dirus in the late Irvingtonian, probably in more open terrain in the midcontinent, and later extending its way to the east and displacing Canis armbrusteri.[19]:181 The timing of Canis dirus would therefore be the Late Irvingtonian of California and Nebraska, and the Rancholabrean of Canada, the United States, Mexico, Venezuela, Ecuador, Bolivia and Peru.[19]:146
The discovery of a fossil in the Horse Room of the Salamander Cave in the Black Hills of South Dakota may possibly be that of a Dire wolf, and if so then it is also one of the earliest records.[13] The specimen was catalogued as Canis cf. C. dirus[21] (where cf. in Latin means confer, uncertain). A horse fossil found in the room gave a uranium-series dating of 252,000 years before present (YBP) and the Canis cf. dirus specimen was assumed to be from the same period.[21][13] If the material actually represents Canis dirus, it would be a late Irvingtonian record of the species.[22]
South America

Canis dirus made its appearance in South America in the late Pleistocene, and seems to have been restricted to the north and west coasts. Its remains have not been found in Argentina that produced Canis gezi and Canis nehringi, and their remains have not been found elsewhere in South America.[19]:148 Some researchers have proposed that Canis dirus may have originated in South America.[18]:116[13][24] In 1988 a study of these two large South American wolves described them, with Canis gezi found in South American Ensenadan deposits that relate to the North American late Blancan and Irvingtonian, and Canis nehringi found in South American Lujanian deposits that relate to the Late Pleistocene. Given their similarities and timeframes, it was proposed that Canis gezi was the ancestor of Canis nehringi.[8]:113 The study indicated that Canis gezi was most similar to the late Irvingtonian Aenocyon dirus nebrascensis and was its sister taxa,[8]:55 but Canis nehringi had a closer relationship to Rancholabrean Canis dirus.[8]:113 The study found that Canis dirus was the most derived genus Canis species in the New World, and compared to Canis nehringi was larger in size and construction of its lower molars that were increased for more efficient predation.[8]:113 In 2009, Tedford proposed that because there was now seen a link between Canis armbrusteri and the Rancholabrean Canis dirus, that a case could be argued for a collateral South American lineage linking Canis gezi with Canis nehringi. These two clades share dental and cranial similarities developed for hypercarnivory, suggesting a common ancestor for both clades.[19]:148
In 2010, a study found that DNA analysis and the dental characteristics of South America hypercanivorous canids showed a "South American clade" and the Canis clade. Canis gezi was a member of the "South American clade" of carnivores, but Canis dirus and Canis nehringi were included as derived species in the Canis clade. Canis dirus was the sister taxon of Canis lupus, but the scientific scoring used in this study for Canis nehringi was identical to those observed in Canis dirus, which supports the proposal that both could have been the same species.[25]:472
Radiocarbon dating
The age of most dire wolf localities is determined solely by biostratigraphy, although some sites have been radiocarbon dated. A specimen from Powder Mill Creek Cave, Missouri, was dated at 13,170 YBP.[13] Canis dirus and Smilodon fatalis are the two most common carnivorans from Rancho La Brea and many have been radiocarbon dated, with the most recent Canis dirus specimen dated at 11,384 YBP and 4 of the oldest dated at 28,230 YBP.[26]
Description
The dire wolf averaged about 1.5 m (4.9 ft) in length and weighed between 50 and 79 kg (110 and 174 lb),[27][28] which made it the largest species in the genus Canis.[29] Limb elements are rarely found outside the La Brea Tar Pits, which makes it hard to compare the size of average individuals between populations. The dire wolf is estimated to have been 8% shorter at the shoulder than the modern Northwestern wolf, and of equal height to the typical gray wolf, but more heavily built.[30] With the exception of the canine teeth in some populations, male and female body and teeth sizes evidence no major sexual dimorphism, similar to most canines. In some populations, males’ canine teeth were considerably larger, suggesting male competition for breeding access. In other populations, lack of dimorphism in the canine teeth suggests little competition.[31]
Despite superficial similarities to the gray wolf, the two species differed significantly. Today’s largest gray wolves would have been of similar size to an average dire wolf; the largest dire wolves would have been considerably larger than any modern gray wolf. The dire wolf is calculated to have weighed 25% more than living gray wolves.[30]
Many of these characteristics were needed to fight off and prey on larger megafauna.[32] The legs of the dire wolf were proportionally shorter and sturdier than those of the gray wolf, and its brain case was smaller than that of a similarly sized gray wolf.[33]
Skull and dentition
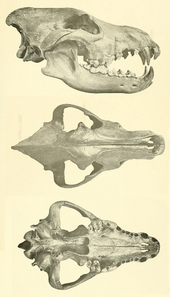
The dire wolf's teeth were similar to the gray wolf's, only slightly larger, pointing to a hypercarnivorous to mesocarnivorous activity. The dietary characteristics were primarily carnivorous, as well as partially omnivorous.[35] Other than size, there is no significant special adaptations in the mandibular corpus of the dire wolf to set it apart from the gray wolf.[36]
The dire wolf was characterized by a relatively larger temporalis muscle that was capable of generating more force than that in the extant gray wolf.[37] The data from one study found that the biteforce at the canine tooth was 893 N for the dire wolf and 593 N for the extant gray wolf, and the biteforce per kilogram of body mass measured 163 for the dire wolf and 136 for the extant gray wolf.[38]
The slicing teeth (P4, the carnassial) on the upper jaw of C. dirus are larger than those of the gray wolf, but those on the lower jaw are similar. The temporalis of the dire wolf could generate more force than seen in modern gray wolves, suggesting stronger killing bites.[39]
Many paleontologists have proposed that the dire wolf may have used its relatively large teeth to crush bone, an idea supported by the frequency of large amounts of wear on the crowns of their fossilized teeth. The upper carnassial had much larger blade than that of the gray wolf, indicating greater slicing ability. It had a longer temporal fossa and broader zygomatic arches, indicating the presence of a large temporalis muscle capable of generating slightly more force than a gray wolf's.[40] However, other scientists have noted the dorsoventral and labiolingual force profiles are indistinguishable from those of other canids, such as gray wolves, coyotes and African wild dogs, which refutes the claim that it was a bone-cracking ecomorph.[41]
Dire wolf teeth lacked the craniodental adaptations of habitual bonecrushers such as hyenas and borophagines.[40] The dire wolf's robust skull and susceptibility to tooth breakage suggests that it hunted for large prey or scavenged the carcasses of large prey.[42] The dorsoventrally weak symphyseal region indicates it killed in a manner similar to its modern relatives, by delivering a series of shallow bites, strongly indicating pack hunting behaviour. However, the incidence of broken postcarnassial molars is much higher than in fossil gray wolves, indicating the species was probably less adapted to bone crushing when compared to the gray wolf.[41]
Behavior and ecology

Dire wolves' overpowering bite, 129% of the force of the modern gray wolf, could hold and subdue their prey. As inferred from their large bodies and carnivorous teeth, they often took on large prey or megafauna, made possible by traveling in packs. Dire wolves were not specialized hunters—they fed on whatever megafauna was abundant.[44]
Compared to modern species, a remarkable number of dire wolf specimens from the La Brea pits showed evidence of having broken their teeth in life. Specimens in the older part of the pit exhibited more tooth wear than those in the younger pit, which could be a result of either the older part of the pit containing more senior dire wolf specimens or a diet that included harder foods such as carcasses and bones.[45][46] Another explanation for increased tooth wear and breakage could be intense competition among carnivores forcing C. dirus to eat as much of the carcass as quickly as possible.[46]
The large amount of remains at the Rancho La Brea tar pits suggest that dire wolves savaged animals trapped there, with some becoming trapped and preserved themselves. It can also be inferred that they were social predators, as those are most successful in defending carcasses.[47]
Habitat and distribution

The habitat of C. dirus varied considerably. In North America, it ranged from plains and grasslands to forested mountain areas. In South America, it occupied areas of arid savannah.[40] The dire wolf lived in several habitats, tropical marsh with thorn-scrub to deciduous forest including some component of nearby grassland, and from sea level up to 2255 m (7400 feet). It was widespread, and its remains have been found in 136 places, from Alberta, Canada, to Tarija, Bolivia. Although the dire wolf was recorded as far north as Alberta, its remains have never been found at higher latitudes. The dire wolf may have migrated to northern Canada during interglacial periods, but its remains would likely have been obliterated by later glacial activity.[13] The largest known dire wolves specimens have also said to have been recovered from Florida. So even though the dire wolf has migrated it may have found to be seasonal.[29]
The dire wolf has an extremely extensive fossil range. It is known to be located as far north as Canada and south into southern Bolivia. Ten localities in Mexico are known to contain Canis dirus: Cedazo, Aguascalientes; Comondu, Baja California; El Cedral, San Luis Potosí; El Tajo Quarry, Tequixquiac, and Valsequillo, all in Distrito Federal, Mexico; Lago de Chapala, Jalisco; Loltun Cave, Yucatán; Potrecito, Sinaloa; and San Josecito Cave, Nuevo León. The majority of these localities occur in the central and southeast-central portions of Mexico with few occurrences in the north or west. Of the central localities, San Josecito Cave and Cedazo have the greatest number of individuals of Canis dirus collected from a single locality; other localities in Mexico are known by only a few specimens.[48]
The dire wolf is well known for its unusually high representation in La Brea Tar Pits in California. Over 200,000 fragments representing more than 4,000 individual dire wolves have been recovered from the tar pits, more than any other mammal species.[30] This large number suggests the dire wolf, like modern wolves and dogs, hunted in packs. The abundance of remains of the gray wolf (C. lupus, also known as C. furlongi) in the tar pits is about 1% that of the dire wolf.[49]
If Canis dirus originated in North America, the species likely dispersed into South America via the Andean corridor,[8][13] a proposed pathway for temperate mammals to migrate from Central to South America because of the favorable cool, dry and open habitats that characterized the region at times. This most likely happened during a glacial period, however, as the pathway then consisted of open, arid regions and savanna whereas during inter-glacial periods, it would have been characterized by tropical rain forest habitat.[50][13]
Competitors
During the Late Pleistocene (125,000 years ago) the gray wolf (C. lupus) crossed into North America on the Bering land bridge and competed with the dire wolf. Overlapping fossil findings of the extinct saber tooth cat, the Smilodon, shows that the dire wolf had these as competition in North America.[51] Both species were social animals that hunt in packs and preyed on the same animals.[51]
Extinction


The dire wolf was one of the abundant Pleistocene megafauna. Starting about 16,000 years ago, coinciding with the end of the last glacial period and the arrival of humans in North America, most of the large mammals upon which the dire wolf depended for prey began to die out, possibly as a result of climate and/or human-induced changes.[53] Approximately 10,000 years ago the dire wolf became extinct along with most other American megafauna.[54][55] Their demise, along with that of other large Pleistocene carnivores, was found to be related to the extinction of megafaunal prey.[42]
See also
References
- 1 2 Leidy, J. 1858: Notice of remains of extinct vertebrata, from the Valley of the Niobrara River, collected during the Exploring Expedition of 1857, in Nebraska, under the command of Lieut. G. K. Warren, U. S. Top. Eng., by Dr. F. V. Hayden, Geologist to the Expedition, Proceedings: Academy of Natural Sciences of Philadelphia, volume 10:1858 p21. LINK:
- 1 2 Leidy, J. 1854: Notice of some fossil bones discovered by Mr. Francis A. Lincke, in the banks of the Ohio River, Indiana. Proceedings: Academy of Natural Sciences of Philadelphia 7, p200 LINK:
- 1 2 Leidy, J. 1869. The extinct mammalian fauna of Dakota and Nebraska, including an account of some allied forms from other localities, together with a synopsis of the mammalian remains of North America. Journal of the Academy of Natural Sciences of Philadelphia 7: p368 LINK:
- 1 2 Allen, J. A. LINK: (1876). "Description of some remains of an extinct species of wolf and an extinct species of deer from the lead region of the upper Mississippi". American Journal of Science (61): 47.
- ↑ Sellards, E.H. 1916. Human remains and associated fossils from the Pleistocene of Florida. Annual Report of the Florida Geological Survey 8: p152 LINK:
- 1 2 Merriam, J.C. 1918. Note on the systematic position of the wolves of the Canis dirus group. Bulletin of the Department of Geology of the University of California 10: p533 LINK:
- 1 2 Frick, C. 1930. Alaska’s frozen fauna. Natural History 30: 71–80.
- 1 2 3 4 5 6 Berta, A. 1988. Quaternary evolution and biogeography of the large South American Canidae (Mammalia: Carnivora). University of California Publiations in Geological Sciences 132:1–49.
- ↑ Page Museum. "Page Museum—La Brea Tar Pits". The Natural History Museum of Los Angeles County Foundation.
- ↑ Fossilworks website Canis dirus full listing
- ↑ Kurten, B. 1984: Geographic differentiation in the Rancholabrean dire wolf (Canis dirus Leidy) in North America. In Genoways, H. H. & Dawson, M. R. (eds.): Contributions in Quaternary Vertebrate Paleontology: A Volume in Memorial to John E. Guilday, 218–227. Carnegie Museum of Natural History Special Publication 8
- ↑ WANG, X. 1990. Pleistocene dire wolf remains from the Kansas River with notes on dire wolves in Kansas. Occasional Papers of the Museum of Natural History, University of Kansas 137:1–7.
- 1 2 3 4 5 6 7 8 Dundas, R.G. (1999). "Quaternary records of the dire wolf, Canis dirus, in North and South America" (PDF). Boreas 28 (3): 375–385. doi:10.1111/j.1502-3885.1999.tb00227.x.
- ↑ Troxell, E. L. (1915). American Journal of Science 189: 613–618. LINK:
- ↑ Marc Stevenson, 1978, Wolf and Man: Evolution in Parallel, edited by Roberta L. Hall, Henry S. Sharp, Academic Press Inc. New York, Chapter 9, p180 LINK:
- ↑ Lundelius, E. L. 1972. Fossil vertebrates, late Pleistocene Ingleside Fauna, San Patricio County, Texas. Bureau of Economic Geology Report of Investigations no.77
- ↑ Fossilworks website Lundelius 1972
- 1 2 3 R. M. Nowak. 1979. North American Quaternary Canis. Monograph of the Museum of Natural History, University of Kansas 6:1-154, page 106 LINK:
- 1 2 3 4 5 6 7 8 9 10 Tedford, Richard H.; Wang, Xiaoming; Taylor, Beryl E. (2009). "Phylogenetic Systematics of the North American Fossil Caninae (Carnivora: Canidae)" (PDF). Bulletin of the American Museum of Natural History 325: 1–218. doi:10.1206/574.1.
- ↑ Flynn, John J. and Gina D. Wesley-Hunt. 2005. "Phylogeny of the Carnivora: Basal Relationships Among the Carnivoramorphans, and Assessment of the Position of 'Miacoidea' Relative to Carnivora." Journal of Systematic Paleontology, 3: 1-28.
- 1 2 Mead, J. I., Manganaro, C., Repenning, C. A. & Agenbroad, L. D. 1996: Early Rancholabrean mammals from Salamander Cave, Black Hills, South Dakota. In Stewart, K. M. & Seymour, K. L. (eds.): Palaeoecology and Palaeoenvironments of Late Cenozoic Mammals, Tributes to the Career of C. S. (Rufus) Churcher, 458–482. University of Toronto Press, Toronto, Ontario, Canada.
- ↑ Lundelius, E.L. and Bell C.J., Late Cretaceous and Cenozoic Mammals of North America: Biostratigraphy and Geochronology, edited by Michael O. Woodburne Columbia university press 2004, Ch7 p285. LINK:
- ↑ Artwork by Sergio De la Rosa Martinez (refer to the Summary of this pix)
- ↑ B. Kurten and E. Anderson. 1980. Pleistocene mammals of North America 1-442
- ↑ Prevosti, Francisco J. (2010). "Phylogeny of the large extinct South American Canids (Mammalia, Carnivora, Canidae) using a "total evidence" approach". Cladistics 26 (5): 456–481. doi:10.1111/j.1096-0031.2009.00298.x.
- ↑ O'Keefe FR,Fet EV,Harris JM. 2009. Compilation, calibration, and synthesis of faunal and floral radiocarbon dates, Rancho La Brea, California. Contrib Sci 518: 1–16 refer Table 2
- ↑ "Wolves, Coyotes, and Dogs (Genus Canis)". Museum.state.il.us. Retrieved 2011-10-23.
- ↑ "San Diego Natural History Museum Fossil Mysteries Field Guide: Dire Wolf". Sdnhm.org. Retrieved 2011-10-23.
- 1 2 Canis dirus. Fossil Species of Florida. Florida Museum of Natural History.
- 1 2 3 Anyonge, W.; Roman, C. (2006). "New body mass estimates for Canis dirus, the extinct Pleistocene dire wolf". Journal of Vertebrate Paleontology 26: 209–212. doi:10.1671/0272-4634(2006)26[209:NBMEFC]2.0.CO;2.
- ↑ Van Valkenburgh, B.; Sacco, T. (2002). "Sexual dimorphism, social behavior, and intrasexual competition in large Pleistocene carnivorans". Journal of Vertebrate Paleontology 22: 164–169. doi:10.1671/0272-4634(2002)022[0164:SDSBAI]2.0.CO;2.
- ↑ Hodnett, John-Paul; Mead, Jim I.; Baez, A. (2009). "Dire Wolf, Canis dirus (mammalia; Carnivora; Canidae), from the Late Pleistocene (Rancholabrean) of East-Central Sonora, Mexico". The Southwest Naturalist 54 (1): 74–81. doi:10.1894/CLG-12.1. Retrieved 31 October 2012.
- ↑ "Wolves, Coyotes, and Dogs (Genus Canis)". Illinois State Museum. Retrieved 2010-06-21.
- ↑ Merriam, J.C. (1911). The fauna of Rancho La Brea 1. University of California - Berkeley. pp. 224–225.
- ↑ Nowak, R. M. (1991). Walker's Mammals of the World. Maryland, Johns Hopkins University Press (edited volume) II
- ↑ Anyonge, W.; Baker A. (2006). "Craniofacial morphology and feeding behavior in Canis dirus, the extinct Pleistocene dire wolf". Journal of Zoology 269 (3): 309–316. doi:10.1111/j.1469-7998.2006.00043.x.
- ↑ Anyonge, W.; Baker, A. (2006). "Craniofacial morphology and feeding behavior in Canis dirus, the extinct Pleistocene dire wolf". Journal of Zoology 269 (3): 309–316. doi:10.1111/j.1469-7998.2006.00043.x.
- ↑ Wroe, S.; McHenry, C.; Thomason, J. (2005). "Bite club: Comparative bite force in big biting mammals and the prediction of predatory behaviour in fossil taxa". Proceedings of the Royal Society B: Biological Sciences 272 (1563): 619. doi:10.1098/rspb.2004.2986.
- ↑ "Dire Wolf Fact Sheet". San Diego Zoo. Retrieved 31 October 2012.
- 1 2 3 Anyonge, William; Baker, A. (2006). "Craniofacial morphology and feeding behavior in C. dirus, the extinct Pleistocene dire wolf". Journal of Zoology 269 (3): 309–316. doi:10.1111/j.1469-7998.2006.00043.x.
- 1 2 Therrien, F. (2005). "Mandibular force profiles of extant carnivorans and implications for the feeding behaviour of extinct predators". Journal of Zoology 267 (3): 249. doi:10.1017/S0952836905007430.
- 1 2 Fox-Dobbs, K; Bump, J.K.; Peterson, R.O.; Fox, D.L.; Koch, P.L. (2007). "Carnivore-specific stable isotope variables and variation in the foraging ecology of modern and ancient wolf populations: case studies from Isle Royale, Minnesota, and La Brea" (PDF). Canadian Journal of Zoology 85 (4): 458–471. doi:10.1139/Z07-018. Retrieved 24 October 2013.
- ↑ Rancho la Brea Tar Pool. Restoration by Bruce Horsfall for W.B. Scott.
- ↑ Wroe, S.; McHenry, C.; Thomason, J. (2005). "Bite club: Comparative bite force in big biting mammals and the prediction of predatory behaviour in fossil taxa". Proceedings of the Royal Society B: Biological Sciences 272 (1563): 619–25. doi:10.1098/rspb.2004.2986. PMC 1564077. PMID 15817436.
- ↑ Binder, W. J.; Thompson, E. N.; Van Valkenburgh, B. (2002). "Temporal variation in tooth fracture among Rancho La Brea dire wolves". Journal of Vertebrate Paleontology 22 (2): 423–428. doi:10.1671/0272-4634(2002)022[0423:TVITFA]2.0.CO;2.
- 1 2 Binder, Wendy J.; Van Valkenburgh, Blaire (28 Jan 2010). "A comparison of tooth wear and breakage in Rancho La Brea sabertooth cats and dire wolves across time". Journal of Vertebrate Paleontology 30 (1): 255–261. doi:10.1080/02724630903413016.
- ↑ McHorse, Brianna K.; Orcutt John D., Davis Edward B.; Davis, Edward B. (15 April 2012). "The carnivoran fauna of Rancho La Brea: Average or aberrant?". Palaeogeography, Palaeoclimatology, Palaeoecology. 329–330: 118–123. doi:10.1016/j.palaeo.2012.02.022.
- ↑ Hodnett, John-Paul; Mead Jim; Baez, A. (March 2009). "Dire Wolf, Canis dirus (Mammalia; Carnivora; Canidae), from the Late Pleistocene (Rancholabrean) of East-Central Sonora, Mexico". The Southwestern Naturalist 54.1: 74–81. doi:10.1894/CLG-12.1.
- ↑ Stock, C. (November 1929). "A Census of the Pleistocene Mammals of Rancho La Brea, Based on the Collections of the Los Angeles Museum". Journal of Mammalogy 10 (4): 281–289. doi:10.2307/1374112. JSTOR 1374112.
- ↑ Webb, S. D. 1991: Ecogeography and the great American interchange. Paleobiology 17(3), 266–280
- 1 2 Carbone, C.; Maddox, T.; Funston, P. J.; Mills, M. G. L.; Grether, G. F.; Van Valkenburgh, B. (2009). "Parallels between playbacks and Pleistocene tar seeps suggest sociality in an extinct sabretooth cat, Smilodon". Biology Letters 5: 81–5. doi:10.1098/rsbl.2008.0526. PMC 2657756. PMID 18957359.
- ↑ Rancho la Brea. Restoration by Chas. R. Knight. Mural for Amer. Museum Hall of Man. Coast Range in back ground, Old Baldy at left
- ↑ Prehistoric Predators. National Geographic (DVD ASIN-B00120TJFE). February 12, 2008.
- ↑ Zabludoff, Marc (2009). Dire Wolf. Marshall Cavendish. pp. 24–27. ISBN 978-0-7614-3998-1.
- ↑ Fiedal, Stuart (2009). "Sudden Deaths: The Chronology of Terminal Pleistocene Megafaunal Extinction". In Haynes, Gary. American Megafaunal Extinctions at the End of the Pleistocene. Springer. pp. 21–37. doi:10.1007/978-1-4020-8793-6_2. ISBN 978-1-4020-8792-9.
{kind=link}
{kind=link}
External links
| Wikispecies has information related to: Canis dirus |
| Wikimedia Commons has media related to Canis dirus. |
| ||||||||||||||||||||||||||