Antifreeze protein
| Insect antifreeze protein | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Structure of the Tenebrio molitor beta-helical antifreeze protein[1] | |||||||||
| Identifiers | |||||||||
| Symbol | AFP | ||||||||
| Pfam | PF02420 | ||||||||
| InterPro | IPR003460 | ||||||||
| SCOP | 1ezg | ||||||||
| SUPERFAMILY | 1ezg | ||||||||
| |||||||||
| Choristoneura fumiferana antifreeze protein (CfAFP) | |||||||||
|---|---|---|---|---|---|---|---|---|---|
 Structure of Choristoneura fumiferana (spruce budworm) beta-helical antifreeze protein[2] | |||||||||
| Identifiers | |||||||||
| Symbol | CfAFP | ||||||||
| Pfam | PF05264 | ||||||||
| InterPro | IPR007928 | ||||||||
| SCOP | 1m8n | ||||||||
| SUPERFAMILY | 1m8n | ||||||||
| OPM superfamily | 392 | ||||||||
| OPM protein | 1l0s | ||||||||
| |||||||||
Antifreeze proteins (AFPs) or ice structuring proteins (ISPs) refer to a class of polypeptides produced by certain vertebrates, plants, fungi and bacteria that permit their survival in subzero environments. AFPs bind to small ice crystals to inhibit growth and recrystallization of ice that would otherwise be fatal.[3] There is also increasing evidence that AFPs interact with mammalian cell membranes to protect them from cold damage. This work suggests the involvement of AFPs in cold acclimatization.[4]
Non-colligative properties
Unlike the widely used automotive antifreeze, ethylene glycol, AFPs do not lower freezing point in proportion to concentration. Rather, they work in a noncolligative manner. This phenomenon allows them to act as an antifreeze at concentrations 1/300th to 1/500th of those of other dissolved solutes. Their low concentration minimizes their effect on osmotic pressure.[4] The unusual properties of AFPs are attributed to their affinity for specific ice crystal surfaces.[5]
Thermal hysteresis
AFPs create a difference between the melting point and freezing point known as thermal hysteresis. The addition of AFPs at the interface between solid ice and liquid water inhibits the thermodynamically favored growth of the ice crystal. Ice growth is kinetically inhibited by the AFPs covering the water-accessible surfaces of ice.[5]
Thermal hysteresis is easily measured in the lab with a nanolitre osmometer. Organisms differ in their values of thermal hysteresis. The maximum level of thermal hysteresis shown by fish AFP is approximately -1.5 °C (29.3 °F). However, insect antifreeze proteins are 10–30 times more active than fish proteins. This difference probably reflects the lower temperatures encountered by insects on land. In contrast, aquatic organisms are exposed only to –1 to –2 °C below freezing. During the extreme winter months, the spruce budworm resists freezing at temperatures approaching –30 °C.[4] The Alaskan beetle Upis ceramboides can survive in a temperature of –60 °C by using antifreeze agents that are not proteins.[6]
The rate of cooling can influence the thermal hysteresis value of AFPs. Rapid cooling can substantially decrease the nonequilibrium freezing point, and hence the thermal hysteresis value. Consequently, organisms cannot necessarily adapt to their subzero environment if the temperature drops abruptly.[4]
Freeze tolerance versus freeze avoidance
Species containing AFPs may be classified as
Freeze avoidant: These species are able to prevent their body fluids from freezing altogether. Generally, the AFP function may be overcome at extremely cold temperatures, leading to rapid ice growth and death.
Freeze tolerant: These species are able to survive body fluid freezing. Some freeze tolerant species are thought to use AFPs as cryoprotectants to prevent the damage of freezing, but not freezing altogether. The exact mechanism is still unknown. However, it is thought AFPs may inhibit recrystallization and stabilize cell membranes to prevent damage by ice.[7] They may work in conjunction with protein ice nucleators (PINs) to control the rate of ice propagation following freezing.[7]
Diversity
There are many known nonhomologous types of AFPs.
Fish AFPs
Antifreeze glycoproteins or AFGPs are found in Antarctic notothenioids and northern cod. They are 2.6-3.3 kD.[8] AFGPs evolved separately in notothenioids and northern cod. In notothenioids, the AFGP gene arose from an ancestral trypsinogen-like serine protease gene.[9]
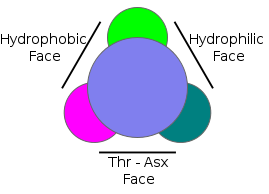
Type I AFP is found in winter flounder, longhorn sculpin and shorthorn sculpin. It is the best documented AFP because it was the first to have its three-dimensional structure determined.[10] Type I AFP consists of a single, long, amphipathic alpha helix, about 3.3-4.5 kD in size. There are three faces to the 3D structure: the hydrophobic, hydrophilic, and Thr-Asx face.[10]
Type I-hyp AFP (where hyp stands for hyperactive) are found in several righteye flounders. It is approximately 32 kD (two 17 kD dimeric molecules). The protein was isolated from the blood plasma of winter flounder. It is considerably better at depressing freezing temperature than most fish AFPs.[11]
Type II AFPs are found in sea raven, smelt and herring. They are cysteine-rich globular proteins containing five disulfide bonds.[12] Type II AFPs likely evolved from calcium dependent (c-type) lectins.[13] Sea ravens, smelt, and herring are quite divergent lineages of teleost. If the AFP gene were present in the most recent common ancestor of these lineages, it's peculiar that the gene is scattered throughout those lineages, present in some orders and absent in others. It has been suggested that lateral gene transfer could be attributed to this discrepancy, such that the smelt acquired the type II AFP gene from the herring.[14]
Type III AFPs are found in Antarctic eelpout. They exhibit similar overall hydrophobicity at ice binding surfaces to type I AFPs. They are approximately 6kD in size.[8] Type III AFPs likely evolved from a sialic acid synthase gene present in Antarctic eelpout. Through a gene duplication event, this gene—which has been shown to exhibit some ice-binding activity of its own—evolved into an effective AFP gene.[15]
Type IV AFPs are found in longhorn sculpins. They are alpha helical proteins rich in glutamate and glutamine.[16] This protein is approximately 12KDa in size and consists of a 4-helix bundle.[16] Its only posttranslational modification is a pyroglutamate residue, a cyclized glutamine residue at its N-terminus.[16] Scientists at the University of Guelph in Canada are currently examining the role of this pyroglutame residue in the antifreeze activity of type IV AFP from the longhorn sculpin.
Plant AFPs
The classification of AFPs became more complicated when antifreeze proteins from plants were discovered.[17] Plant AFPs are rather different from the other AFPs in the following aspects:
- They have much weaker thermal hysteresis activity when compared to other AFPs.[18]
- Their physiological function is likely in inhibiting the recrystallization of ice rather than in the preventing ice formation.[18]
- Most of them are evolved pathogenesis-related proteins, sometimes retaining antifungal properties.[18]
See also dehydrin
Insect AFPs
There are two types of insect antifreeze proteins, Tenebrio and Dendroides AFPs which are both in different insect families. They are similar to one another, both being hyperactive (i.e. greater thermal hysteresis value) and consist of varying numbers of 12- or 13-mer repeats of approximately 8.3 to 12.5 kD. Throughout the length of the protein, at least every sixth residue is a cysteine.[7]
Tenebrio or Type V AFPs are found in beetles,[19] whereas Dendroides or Choristoneura fumiferana AFPs are found in some Lepidoptera.
Sea ice organisms AFPs
AFPs were also found in microorganisms living in sea ice. The diatoms Fragilariopsis cylindrus and F. curta play a key role in polar sea ice communities, dominating the assemblages of both platelet layer and within pack ice. AFPs are widespread in these species, and the presence of AFP genes as a multigene family indicates the importance of this group for the genus Fragilariopsis.[20] AFPs identified in F. cylindrus belong to an AFP family which is represented in different taxa and can be found in other organisms related to sea ice (Colwellia spp., Navicula glaciei, Chaetoceros neogracile and Stephos longipes and Leucosporidium antarcticum[21][22]) and Antarctic inland ice bacteria (Flavobacteriaceae),[23][24] as well as in cold-tolerant fungi (Typhula ishikariensis, Lentinula edodes and Flammulina populicola.[25][26])
Evolution
The remarkable diversity and distribution of AFPs suggest the different types evolved recently in response to sea level glaciation occurring 1-2 million years ago in the Northern hemisphere and 10-30 million years ago in Antarctica. This independent development of similar adaptations is referred to as convergent evolution.[4] There are two reasons why many types of AFPs are able to carry out the same function despite their diversity:
- Although ice is uniformly composed of oxygen and hydrogen, it has many different surfaces exposed for binding. Different types of AFPs may interact with different surfaces.
- Although the five types of AFPs differ in their primary sequence of amino acids, when each folds into a functioning protein, they may share similarities in their three-dimensional or tertiary structure that facilitates the same interactions with ice.[4][27]
{kind=link}
Mechanisms of action
AFPs are thought to inhibit growth by an adsorption–inhibition mechanism.[28] They adsorb to nonbasal planes of ice, inhibiting thermodynamically favored ice growth.[29] The presence of a flat, rigid surface in some AFPs seems to facilitate its interaction with ice via Van der Waals force surface complementarity.[30]
Binding to ice
Normally, ice crystals grown in solution only exhibit the basal (0001) and prism faces (1010), and appear as round and flat discs.[5] However, it appears the presence of AFPs exposes other faces. It now appears the ice surface 2021 is the preferred binding surface, at least for AFP type I.[31] Through studies on type I AFP, ice and AFP were initially thought to interact through hydrogen bonding (Raymond and DeVries, 1977). However, when parts of the protein thought to facilitate this hydrogen bonding were mutated, the hypothesized decrease in antifreeze activity was not observed. Recent data suggest hydrophobic interactions could be the main contributor.[32] It is difficult to discern the exact mechanism of binding because of the complex water-ice interface. Currently, attempts to uncover the precise mechanism are being made through use of molecular modelling programs (molecular dynamics or the Monte Carlo method).[3][5]
Binding mechanism and antifreeze function
According to the structure and function study on the antifreeze protein from the fish winter flounder,[33] the antifreeze mechanism of the type-I AFP molecule was shown to be due to the binding to an ice nucleation structure in a zipper-like fashion through hydrogen bonding of the hydroxyl groups of its four Thr residues to the oxygens along the ![[01\overline{1}2]](../I/m/5081bd37778f6a7c976ffbc9f5a1c885.png) direction in ice lattice, subsequently stopping or retarding the growth of ice pyramidal planes so as to depress the freeze point.[33]
direction in ice lattice, subsequently stopping or retarding the growth of ice pyramidal planes so as to depress the freeze point.[33]
The above mechanism can be used to elucidate the structure-function relationship of other antifreeze proteins with the following two common features:
- recurrence of a Thr residue (or any other polar amino acid residue whose side-chain can form a hydrogen bond with water) in an 11-amino-acid period along the sequence concerned, and
- a high percentage of an Ala residue component therein.[33]
History
In the 1950s, Norwegian scientist Scholander set out to explain how Arctic fish can survive in water colder than the freezing point of their blood. His experiments led him to believe there was “antifreeze” in the blood of Arctic fish.[3] Then in the late 1960s, animal biologist Arthur DeVries was able to isolate the antifreeze protein through his investigation of Antarctic fish.[34] These proteins were later called antifreeze glycoproteins (AFGPs) or antifreeze glycopeptides to distinguish them from newly discovered nonglycoprotein biological antifreeze agents (AFPs). DeVries worked with Robert Feeney (1970) to characterize the chemical and physical properties of antifreeze proteins.[35] In 1992, Griffith et al. documented their discovery of AFP in winter rye leaves. Around the same time, Urrutia, Duman and Knight (1992) documented thermal hysteresis protein in angiosperms. The next year, Duman and Olsen noted AFPs had also been discovered in over 23 species of angiosperms, including ones eaten by humans.[36] As well, they reported their presence in fungi and bacteria.
Name change
Recent attempts have been made to relabel antifreeze proteins as ice structuring proteins to more accurately represent their function and to dispose of any assumed negative relation between AFPs and automotive antifreeze, ethylene glycol. These two things are completely separate entities, and show loose similarity only in their function.[37]
Commercial applications
Commercially, there appear to be countless applications for antifreeze proteins.[38] Numerous fields would be able to benefit from the protection of tissue damage by freezing. Businesses are currently investigating the use of these proteins in:
- increasing freeze tolerance of crop plants and extending the harvest season in cooler climates
- improving farm fish production in cooler climates
- lengthening shelf life of frozen foods
- improving cryosurgery
- enhancing preservation of tissues for transplant or transfusion in medicine[39]
- therapy for hypothermia
Recent news
One recent, successful business endeavor has been the introduction of AFPs into ice cream and yogurt products. This ingredient, labelled ice-structuring protein, has been approved by the Food and Drug Administration. The proteins are isolated from fish and replicated, on a larger scale, in genetically modified yeast.[40]
There is concern from organizations opposed to genetically modified organisms (GMOs), arguing modified antifreeze proteins may cause inflammation.[41] Intake of non genetically modified AFPs in diet is likely substantial in most northerly and temperate regions already.[8] Given the known historic consumption of AFPs, it is safe to conclude their functional properties do not impart any toxicologic or allergenic effects in humans.[8]
As well, the transgenic process of ISP production is widely used in society already. Insulin and rennet are produced using this technology. The process does not impact the product; it merely makes production more efficient and prevents the death of fish which would otherwise be killed to extract the protein.
Currently, Unilever incorporates AFPs into some of its American products, including some popsicles and a new line of Breyers Light Double Churned ice cream bars. In ice cream, AFPs allow the production of very creamy, dense, reduced fat ice cream with fewer additives.[42] They control ice crystal growth brought on by thawing on the loading dock or kitchen table which drastically reduces texture quality.[43]
In November 2009, the Proceedings of the National Academy of Sciences published the discovery of a molecule in an Alaskan beetle that behaves like AFPs, but is composed of saccharides and fatty acids.[6]
A 2010 study demonstrated the stability of superheated water ice crystals in an AFP solution, showing while the proteins can inhibit freezing, they can also inhibit melting.[44]
References
- ↑ Daley ME, Spyracopoulos L, Jia Z, Davies PL, Sykes BD (April 2002). "Structure and dynamics of a beta-helical antifreeze protein". Biochemistry 41 (17): 5515–25. doi:10.1021/bi0121252. PMID 11969412.
- ↑ Leinala EK, Davies PL, Doucet D, Tyshenko MG, Walker VK, Jia Z (September 2002). "A beta-helical antifreeze protein isoform with increased activity. Structural and functional insights". J. Biol. Chem. 277 (36): 33349–52. doi:10.1074/jbc.M205575200. PMID 12105229.
- 1 2 3 http://www.rcsb.org/pdb/101/motm.do?momID=120
- 1 2 3 4 5 6 Fletcher GL, Hew CL, Davies PL (2001). "Antifreeze proteins of teleost fishes". Annu. Rev. Physiol. 63: 359–90. doi:10.1146/annurev.physiol.63.1.359. PMID 11181960.
- 1 2 3 4 Jorov A, Zhorov BS, Yang DS (June 2004). "Study of interaction of winter flounder antifreeze protein with ice". Protein Sci. 13 (6): 1524–37. doi:10.1110/ps.04641104. PMC 2279984. PMID 15152087.
- 1 2 Walters KR Jr, Serianni AS, Sformo T, Barnes BM, Duman JG (2009). "A nonprotein thermal hysteresis-producing xylomannan antifreeze in the freeze-tolerant Alaskan beetle Upis ceramboides". PNAS 106 (48): 20210–5. doi:10.1073/pnas.0909872106. PMC 2787118. PMID 19934038.
- 1 2 3 Duman JG (2001). "Antifreeze and ice nucleator proteins in terrestrial arthropods". Annu. Rev. Physiol. 63: 327–57. doi:10.1146/annurev.physiol.63.1.327. PMID 11181959.
- 1 2 3 4 Crevel RW, Fedyk JK, Spurgeon MJ (July 2002). "Antifreeze proteins: characteristics, occurrence and human exposure". Food Chem. Toxicol. 40 (7): 899–903. doi:10.1016/S0278-6915(02)00042-X. PMID 12065210.
- ↑ Chen; et al. (1997). "Evolution of antifreeze glycoprotein gene from a trypsinogen gene in Antarctic notothenioid fish". PNAS 94 (8): 3811–3816. doi:10.1073/pnas.94.8.3811. PMC 20523. PMID 9108060.
- 1 2 Duman JG, de Vries AL (1976). "Isolation, characterization, and physical properties of protein antifreezes from the winter flounder, Pseudopleuronectes americanus". Comp. Biochem. Physiol., B 54 (3): 375–80. doi:10.1016/0305-0491(76)90260-1. PMID 1277804.
- ↑ Scotter AJ, Marshall CB, Graham LA, Gilbert JA, Garnham CP, Davies PL (October 2006). "The basis for hyperactivity of antifreeze proteins". Cryobiology 53 (2): 229–39. doi:10.1016/j.cryobiol.2006.06.006. PMID 16887111.
- ↑ Ng NF, Hew CL (August 1992). "Structure of an antifreeze polypeptide from the sea raven. Disulfide bonds and similarity to lectin-binding proteins". J. Biol. Chem. 267 (23): 16069–75. PMID 1644794.
- ↑ Ewart; et al. (1992). "Structural and functional similarity between fish antifreeze proteins and calcium-dependent lectins". Biochem Biophys Res Commun 185: 335–340. doi:10.1016/s0006-291x(05)90005-3.
- ↑ Graham; et al. (2008). "Lateral transfer of a lectin-like antifreeze protein gene in fishes". PLOS ONE 3: e2616. doi:10.1371/journal.pone.0002616.

- ↑ Kelley; et al. (2010). "Functional diversification and evolution of antifreeze proteins in the Antarctic fish Lycodichthys dearborni". Journal of Molecular Evolution 71: 111–118. doi:10.1007/s00239-010-9367-6.
- 1 2 3 Deng G, Andrews DW, Laursen RA (January 1997). "Amino acid sequence of a new type of antifreeze protein, from the longhorn sculpin Myoxocephalus octodecimspinosis". FEBS Lett. 402 (1): 17–20. doi:10.1016/S0014-5793(96)01466-4. PMID 9013849.
- ↑ Griffith M, Ala P, Yang DS, Hon WC, Moffatt BA (October 1992). "Antifreeze protein produced endogenously in winter rye leaves". Plant Physiol. 100 (2): 593–6. doi:10.1104/pp.100.2.593. PMC 1075599. PMID 16653033.
- 1 2 3 Griffith M, Yaish MW (August 2004). "Antifreeze proteins in overwintering plants: a tale of two activities". Trends Plant Sci. 9 (8): 399–405. doi:10.1016/j.tplants.2004.06.007. PMID 15358271.
- ↑ Graham LA, Liou YC, Walker VK, Davies PL (August 1997). "Hyperactive antifreeze protein from beetles". Nature 388 (6644): 727–8. doi:10.1038/41908. PMID 9285581.
- ↑ Bayer-Giraldi M, Uhlig C, John U, Mock T, Valentin K (April 2010). "Antifreeze proteins in polar sea ice diatoms: diversity and gene expression in the genus Fragilariopsis". Environ. Microbiol. 12 (4): 1041–52. doi:10.1111/j.1462-2920.2009.02149.x. PMID 20105220.
- ↑ Raymond JA, Fritsen C, Shen K (August 2007). "An ice-binding protein from an Antarctic sea ice bacterium". FEMS Microbiol. Ecol. 61 (2): 214–21. doi:10.1111/j.1574-6941.2007.00345.x. PMID 17651136.
- ↑ Kiko, R. (2010): Acquisition of freeze protection in a sea-ice crustacean through horizontal gene transfer? Polar Biology (33) 543-556.
- ↑ Raymond JA, Christner BC, Schuster SC (September 2008). "A bacterial ice-binding protein from the Vostok ice core". Extremophiles 12 (5): 713–7. doi:10.1007/s00792-008-0178-2. PMID 18622572.
- ↑ http://pnwfungi.org/pdf_files/manuscripts_volume_5/naf20105/naf2010514.pdf
- ↑ Hoshino, T., Kiriaki, M., Ohgiya, S., Fujiwara, M., Kondo, H., Nishimiya, Y., et al. (2003) Antifreeze proteins from snow mold fungi. Can J Bot 81: 1175–1181.
- ↑ Raymond JA, Janech MG (April 2009). "Ice-binding proteins from enoki and shiitake mushrooms". Cryobiology 58 (2): 151–6. doi:10.1016/j.cryobiol.2008.11.009. PMID 19121299.
- ↑ Chen L, DeVries AL, Cheng CH (April 1997). "Convergent evolution of antifreeze glycoproteins in Antarctic notothenioid fish and Arctic cod". Proc. Natl. Acad. Sci. U.S.A. 94 (8): 3817–22. doi:10.1073/pnas.94.8.3817. PMC 20524. PMID 9108061.
- ↑ Raymond JA, DeVries AL (June 1977). "Adsorption inhibition as a mechanism of freezing resistance in polar fishes". Proc. Natl. Acad. Sci. U.S.A. 74 (6): 2589–93. doi:10.1073/pnas.74.6.2589. PMC 432219. PMID 267952.
- ↑ Raymond JA, Wilson P, DeVries AL (February 1989). "Inhibition of growth of non basal planes in ice by fish antifreezes". Proc. Natl. Acad. Sci. U.S.A. 86 (3): 881–5. doi:10.1073/pnas.86.3.881. PMC 286582. PMID 2915983.
- ↑ Yang DS, Hon WC, Bubanko S, Xue Y, Seetharaman J, Hew CL, Sicheri F (May 1998). "Identification of the ice-binding surface on a type III antifreeze protein with a "flatness function" algorithm". Biophys. J. 74 (5): 2142–51. doi:10.1016/S0006-3495(98)77923-8. PMC 1299557. PMID 9591641.
- ↑ Knight CA, Cheng CC, DeVries AL (February 1991). "Adsorption of alpha-helical antifreeze peptides on specific ice crystal surface planes". Biophys. J. 59 (2): 409–18. doi:10.1016/S0006-3495(91)82234-2. PMC 1281157. PMID 2009357.
- ↑ Haymet AD, Ward LG, Harding MM, Knight CA (July 1998). "Valine substituted winter flounder 'antifreeze': preservation of ice growth hysteresis". FEBS Lett. 430 (3): 301–6. doi:10.1016/S0014-5793(98)00652-8. PMID 9688560.
- 1 2 3 Chou KC (January 1992). "Energy-optimized structure of antifreeze protein and its binding mechanism". J. Mol. Biol. 223 (2): 509–17. doi:10.1016/0022-2836(92)90666-8. PMID 1738160.
- ↑ DeVries AL, Wohlschlag DE (March 1969). "Freezing resistance in some Antarctic fishes". Science 163 (3871): 1073–5. doi:10.1126/science.163.3871.1073. PMID 5764871.
- ↑ DeVries AL, Komatsu SK, Feeney RE (June 1970). "Chemical and physical properties of freezing point-depressing glycoproteins from Antarctic fishes". J. Biol. Chem. 245 (11): 2901–8. PMID 5488456.
- ↑ Duman JG, Olsen TM (1993). "Thermal hysteresis protein activity in bacteria, fungi and phylogenetically diverse plants". Cryobiology 30 (3): 322–328. doi:10.1006/cryo.1993.1031.
- ↑ Clarke CJ, Buckley SL, Lindner N (2002). "Ice structuring proteins - a new name for antifreeze proteins". Cryo Letters 23 (2): 89–92. PMID 12050776.
- ↑ Antifreeze proteins and their genes: From basic research to business opportunity
- ↑ Science Daily
- ↑ http://www.impactnottingham.com/2014/09/can-ice-cream-be-tasty-and-healthy/
- ↑ Dortch, Eloise. (2006). Fishy GM yeast used to make ice-cream. Network of Concerned Farmers. Retrieved October 09, 2006
- ↑ Creamy, Healthier Ice Cream? What’s the Catch?
- ↑ Regand A, Goff HD (January 2006). "Ice recrystallization inhibition in ice cream as affected by ice structuring proteins from winter wheat grass". J. Dairy Sci. 89 (1): 49–57. doi:10.3168/jds.S0022-0302(06)72068-9. PMID 16357267.
- ↑ Celik, Y; Graham, LA; Mok, YF; Bar, M; Davies, PL; Braslavsky, I (2010). "Superheating of Ice in the Presence of Ice Binding Proteins". Biophysical Journal 98 (3): 245a. doi:10.1016/j.bpj.2009.12.1331. Lay summary – Physorg.com.
Further reading
- Haymett, A.; Ward, L.; Harding, M. (1999). "Winter Flounder 'anti-freeze' proteins: Synthesis and ice growth inhibition of analogues that probe the relative importance of hydrophobic and hydrogen bonding interactions". Journal of the American Chemical Society 121 (5): 941–948. doi:10.1021/ja9801341. ISSN 0002-7863.
- Sicheri, F.; Yang, D. S. (1995). "Ice-binding structure and mechanism of an antifreeze protein from winter flounder". Nature 375 (6530): 427–431. doi:10.1038/375427a0. PMID 7760940.
External links
- Cold, Hard Fact: Fish Antifreeze Produced in Pancreas
- Antifreeze Proteins: Molecule of the Month, by David Goodsell, RCSB Protein Data Bank