Visual search
Visual search is a type of perceptual task requiring attention that typically involves an active scan of the visual environment for a particular object or feature (the target) among other objects or features (the distractors). Visual search can take place either with or without eye movements. The ability to consciously locate an object (target) amongst a complex array of stimuli (distractors) has been extensively studied over the past 40 years. Practical examples of this can be seen in everyday life such as picking out a product on a supermarket shelf, animals searching for food amongst piles of leaves, trying to find your friend in a large crowd of people and playing visual search tasks such as Where's Wally? Many visual search paradigms have used eye movements as a means to measure the degree of attention given to stimuli.[1][2] However, vast research to date suggests that eye movements move independently of attention and therefore is not a reliable method to examine the role of attention. Much of the previous literature on visual search uses reaction time in order to measure the time taken to detect the target amongst its distractors. An example of this could be a green square (target) amongst a set of red circles (distractors).
Search types
Feature search
Feature search (also known as a "disjunctive" or an "efficient" search) as introduced by Treisman and Gelade (1980) [3] is defined as a parallel process. This type of search is used in cases where the target and the distractors are maximally different, i.e. differentiated by a single property such as color, shape, orientation, or size. An example of this would be to pick out a red circle located within a group of black circles. This search type is very efficient, resulting in quick reaction times for correct responses, and is described as having a "pop out" effect. The number of distractors around the target does not affect reaction times for this type of search, as long as they are maximally similar to one another and maximally dissimilar to the target. Thus, the "pop out" effect is as strong with 4 distractors as it is with 40.[4]
Conjunction search
Conjunction search (inefficient search) also described by Treisman and Gelade (1980) [3] occurs when the target and the distractors share similarities in more than one single visual property such as size, colour, orientation and shape. An example of this can be seen if the target is a black horizontal line while the distractors are made up of white horizontal lines and black and white vertical lines. The target therefore shares orientation (horizontal) but not colour with some of the distractors but also shares colour (black) but not orientation with other distractors, The similarities with the distractors make the target harder to identify as there is no 'pop out' effect as seen in feature search. This top-down (user driven) search produces much slower reaction times as it requires the individual to use previously stored knowledge of the target in order to locate it. In this instance, as the number of distractors increases, the search is less efficient. E.g., you would be much slower to detect the target with forty distractor stimuli as opposed to four.
Reaction time slope
It is also possible to determine the role of attention within visual search experiments through measuring the slope of reaction time over the number of distractors present.[5] That is, when high levels of attention are required when looking at a complex array of stimuli (conjunction search), the slope increases as the reaction times increase. Whereas for a simple visual search task (feature search), the slope is flat due to reaction times being fast and less attention is required.
Visual orienting and attention

One obvious way to select visual information is to turn towards it, also known as visual orienting. This may be a movement of the head and/or eyes towards the visual stimulus, called a saccade. Through a process called foveation, the eyes fixate on the object of interest, making the image of the visual stimulus fall on the fovea of the eye, the central part of the retina with the sharpest visual acuity.
There are two types of orienting:
- Exogenous orienting is caused by a sudden change in the periphery. Attention is therefore externally guided by a stimulus, resulting in a reflexive saccade.
- Endogenous orienting results when attention is guided by the goals of the perceiver. Thus, the focus of attention of the perceiver can be manipulated by the demands of a task. A scanning saccade is triggered endogenously for the purpose of exploring the visual environment.

Visual search relies primarily on endogenous orienting because participants have the goal to detect the presence or absence of a specific target object in an array of other distracting objects.
Visual orienting does not necessarily require overt movement, though.[6] It has been shown that people can covertly (without eye movement) shift attention to peripheral stimuli. [7] In the 1970s, it was found that the firing rate of cells in the parietal lobe of monkeys increased in response to stimuli in the receptive field when they attended to peripheral stimuli, even when no eye movements were allowed.[7] These findings indicate that attention plays a critical role in understanding visual search.
Subsequently, competing theories of attention have come to dominate visual search discourse.[8] The environment contains a vast amount of information. We are limited in the amount of information we are able to process at any one time, so it is therefore necessary that we have mechanisms by which extraneous stimuli can be filtered and only relevant information attended to. In the study of attention, psychologists distinguish between preattentitive and attentional processes.[9] Preattentive processes are evenly distributed across all input signals, forming a kind of "low-level" attention. Attentional processes are more selective and can only be applied to specific preattentive input. A large part of the current debate in visual search theory centres on selective attention and what the visual system is capable of achieving without focal attention.[8]
Theory
Feature integration theory (FIT)
A popular explanation for the different reaction times of feature and conjunction searches is the feature integration theory (FIT), introduced by Treisman and Gelade in 1980. This theory proposes that certain visual features are registered early, automatically, and are coded rapidly in parallel across the visual field using preattentive processes.[10] Experiments show that these features include luminance, colour, orientation, motion direction, and velocity, as well as some simple aspects of form.[11] For example, a red X can be quickly found among any number of black Xs and Os because the red X has the discriminative feature of colour and will "pop out". In contrast, this theory also suggests that in order to integrate two or more visual features belonging to the same object, a later process involving integration of information from different brain areas is needed and is coded serially using focal attention. For example, when locating an orange square among blue squares and orange triangles, neither the colour feature "orange" nor the shape feature "square" is sufficient to locate the search target. Instead, one must integrate information of both colour and shape to locate the target.
Evidence that attention and thus later visual processing is needed to integrate two or more features of the same object is shown by the occurrence of illusory conjunctions, or when features do not combine correctly. For example, if a display of a green X and a red O are flashed on a screen so briefly that the later visual process of a serial search with focal attention cannot occur, the observer may report seeing a red X and a green O.
Guided search model
A second main function of preattentive processes is to direct focal attention to the most "promising" information in the visual field.[8] There are two ways in which these processes can be used to direct attention: bottom-up activation (which is stimulus-driven) and top-down activation (which is user-driven). In the guided search model by Jeremy Wolfe,[12] information from top-down and bottom-up processing of the stimulus is used to create a ranking of items in order of their attentional priority. In a visual search, attention will be directed to the item with the highest priority. If that item is rejected, then attention will move on to the next item and the next, and so forth. The guided search theory follows that of parallel search processing.
An activation map is a representation of visual space in which the level of activation at a location reflects the likelihood that the location contains a target. This likelihood is based on preattentive, featural information of the perceiver. According to the guided search model, the initial processing of basic features produces an activation map, with every item in the visual display having its own level of activation. Attention is demanded based on peaks of activation in the activation map in a search for the target.[12] Visual search can proceed efficiently or inefficiently. During efficient search, performance is unaffected by the number of distractor items. The reaction time functions are flat, and the search is assumed to be a parallel search. Thus, in the guided search model, a search is efficient if the target generates the highest, or one of the highest activation peaks. For example, suppose someone is searching for red, horizontal targets. Feature processing would activate all red objects and all horizontal objects. Attention is then directed to items depending on their level of activation, starting with those most activated. This explains why search times are longer when distractors share one or more features with the target stimuli. In contrast, during inefficient search, the reaction time to identify the target increases linearly with the number of distractor items present. According to the guided search model, this is because the peak generated by the target is not one of the highest.[12]
Biological basis
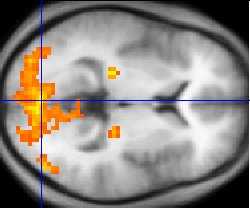
During visual search experiments the posterior parietal cortex has elicited much activation during functional magnetic resonance imaging (fMRI) and electroencephalography (EEG) experiments for inefficient conjunction search, which has also been confirmed through lesion studies. Patients with lesions to the posterior parietal cortex show low accuracy and very slow reaction times during a conjunction search task but have intact feature search remaining to the ipsilesional (the same side of the body as the lesion) side of space. [13] [14] [15] [16] Ashbridge, Walsh, and Cowey in (1997) [17] demonstrated that during the application of transcranial magnetic stimulation (TMS) to the right parietal cortex, conjunction search was impaired by 100 milliseconds after stimulus onset. This was not found during feature search. Nobre, Coull, Walsh and Frith (2003) [18] identified using functional magnetic resonance imaging (fMRI) that the intraparietal sulcus located in the superior parietal cortex was activated specifically to feature search and the binding of individual perceptual features as opposed to conjunction search. Conversely, the authors further identify that for conjunction search, the superior parietal lobe and the right angular gyrus elicit bilaterally during fMRI experiments.
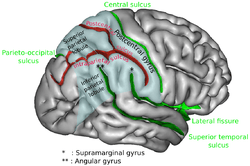
In contrast, Leonards, Sunaert, Vam Hecke and Orban (2000) [19] identified that significant activation is seen during fMRI experiments in the superior frontal sulcus primarily for conjunction search. This research hypothesises that activation in this region may in fact reflect working memory for holding and maintaining stimulus information in mind in order to identify the target. Furthermore, significant frontal activation including the ventrolateral prefrontal cortex bilaterally and the right dorsolateral prefrontal cortex were seen during positron emission tomography for attentional spatial representations during visual search.[20] The same regions associated with spatial attention in the parietal cortex coincide with the regions associated with feature search. Furthermore, the frontal eye field (FEF) located bilaterally in the prefrontal cortex, plays a critical role in saccadic eye movememnts and the control of visual attention [21][22][23]
Moreover, research into monkeys and single cell recording found that the superior colliculus is involved in the selection of the target during visual search as well as the initiation of movements .[24] Conversely, it also suggested that activation in the superior colliculus results from disengaging attention, ensuring that the next stimulus can be internally represented. The ability to directly attend to a particular stimuli during visual search experiments has been linked to the pulvinar nucleus (located in the midbrain) while inhibiting attention to unattended stimuli.[5] Conversely, Bender and Butter (1987) [25] found that during testing on monkeys, no involvement of the pulvinar nucleus was identified during visual search tasks.
Evolution of visual search
There is a variety of speculation about the origin and evolution of visual search in humans. It has been shown that during visual exploration of complex natural scenes, both humans and nonhuman primates make highly stereotyped eye movements.[26] Furthermore, chimpanzees have demonstrated improved performance in visual searches for upright human or dog faces,[27] suggesting that visual search (particularly where the target is a face) is not peculiar to humans and that it may be a primal trait. Research has suggested that effective visual search may have developed as a necessary skill for survival, where being adept at detecting threats and identifying food was essential.

The importance of evolutionarily relevant threat stimuli was demonstrated in a study by LoBue and DeLoache (2008) in which children (and adults) were able to detect snakes more rapidly than other targets amongst distractor stimuli.[28]
Given that the environment in which humans live has changed significantly over time, questions arise as to whether the purpose of visual search is falling away, or whether humans have adapted it to identify new salient targets. Recent research into the relevance of visual search in modern society has include identifying target nutritional information on product labels,[29] identifying salient features while driving[30] and manipulating consumer shopping habits using different shelf display characteristics.[31] Another modern application of visual search has been the development of artificial visual search engines, such as Google Goggles.
Face recognition in visual search
The processing of faces during visual search paradigms has been suggested to take a serial processing form as opposed to an efficient parallel form. Over the past few decades there have been vast amounts of research into face recognition, specifying that faces endure specialized processing within a region called the fusiform face area (FFA) located in the mid fusiform gyrus in the temporal lobe.[32] Debates are ongoing whether both faces and objects are detected and processed in different systems and whether both have category specific regions for recognition and identification.[33][34] Much research to date focuses on the accuracy of the detection and the time taken to detect the face in a complex visual search array. When faces are displayed in isolation, upright faces are processed faster and more accurately than inverted faces,[35][36][37][38] but this effect was observed in non-face objects as well.[39] When faces are to be detected among inverted or jumbled faces, reaction times for intact and upright faces increase as the number of distractors within the array is increased.[40][41][42] Hence, it is argued that the ‘pop out’ theory defined in feature search is not applicable in the recognition of faces in such visual search paradigm. Conversely, the opposite effect has been argued and within a natural environmental scene, the 'pop out' effect of the face is significantly shown. This could be due to evolutionary developments as the need to be able to identify faces that appear threatening to the individual or group is deemed critical in the survival of the fittest.[43] More recently, it was found that faces can be efficeintly detected in a visual search paradigm, if the distracters are non-face objects,[44][45][46] however it is debated whether this apparent 'pop out' effect is driven by a high-level mechanism or by low-level confounding features.[47][48] Furthermore, patients with developmental prosopagnosia, suffering from imparied face identification, generally detect faces normally, suggesting that visual search for faces is facilitated by mechanisms other than the face-identification circuits of the Fusiform Face Area.[49]
Considerations
Ageing
Research indicates that performance in conjunctive visual search tasks significantly improves during childhood and declines in later life.[50] More specifically, young adults have been shown to have faster reaction times on conjunctive visual search tasks than both children and older adults, but their reaction times were similar for feature visual search tasks.[5] This suggests that there is something about the process of integrating visual features or serial searching that is difficult for children and older adults, but not for young adults. Studies have suggested numerous mechanisms involved in this difficulty in children, including peripheral visual acuity,[51] eye movement ability,[52] ability of attentional focal movement,[53] and the ability to divide visual attention among multiple objects.[54]
Studies have suggested similar mechanisms in the difficulty for older adults, such as age related optical changes that influence peripheral acuity,[55] the ability to move attention over the visual field,[56] the ability to disengage attention,[57] and the ability to ignore distractors.[58]
A study by Lorenzo-López et al. (2008) provides neurological evidence for the fact that older adults have slower reaction times during conjunctive searches compared to young adults. Event-related potentials (ERPs) showed longer latencies and lower amplitudes in older subjects than young adults at the P3 component, which is related to activity of the parietal lobes. This suggests the involvement of the parietal lobe function with an age-related decline in the speed of visual search tasks. Results also showed that older adults, when compared to young adults, had significantly less activity in the anterior cingulate cortex and many limbic and occipitotemporal regions that are involved in performing visual search tasks.[59]
Alzheimer's disease
Research has found that people with Alzheimer's disease (AD) are significantly impaired overall in visual search tasks.[60] Surprisingly, AD sufferers manifest enhanced spatial cueing, but this benefit is only obtained for cues with high spatial precision.[61] Abnormal visual attention may underlie certain visuospatial difficulties in patients with (AD). People with AD have hypometabolism and neuropathology in the parietal cortex, and given the role of parietal function for visual attention, patients with AD may have hemispatial neglect, which may result in difficulty with disengaging attention in visual search.[62]
An experiment conducted by Tales et al. (2000) [60] investigated the ability of patients with AD to perform various types of efficient visual search tasks. Their results showed that search rates on the "pop-out" tasks were similar for both AD and control groups, however, people with AD searched significantly slower compared to the control group on the conjunction task. One interpretation of these results is that the visual system of AD patients has a problem with feature binding, such that it is unable to communicate efficiently the different feature descriptions for the stimulus.[60] Binding of features is thought to be mediated by areas in the temporal and parietal cortex, and these areas are known to be affected by AD-related pathology.
Another possibility for the impairment of people with AD on conjunction searches is that there may be some damage to general attentional mechanisms in AD, and therefore any attention-related task will be affected, including visual search.[60]
Tales et al. (2000) detected a double dissociation with their experimental results on AD and visual search. Earlier work was carried out on patients with Parkinson's disease (PD) concerning the impairment patients with PD have on visual search tasks.[63] [64] In those studies, evidence was found of impairment in PD patients on the "pop-out" task, but no evidence was found on the impairment of the conjunction task. As discussed, AD patients show the exact opposite of these results: normal performance was seen on the "pop-out" task, but impairment was found on the conjunction task. This double dissociation provides evidence that PD and AD affect the visual pathway in different ways, and that the pop-out task and the conjunction task are differentially processed within that pathway.
Autism
Studies have consistently shown that autistic individuals performed better and with lower reaction times in feature and conjunctive visual search tasks than matched controls without autism.[65][66] Several explanations for these observations have been suggested. One possibility is that people with autism have enhanced perceptual capacity.[66] This means that autistic individuals are able to process larger amounts of perceptual information, allowing for superior parallel processing and hence faster target location.[67] Second, autistic individuals show superior performance in discrimination tasks between similar stimuli and therefore may have an enhanced ability to differentiate between items in the visual search display.[68] A third suggestion is that autistic individuals may have stronger top-down target excitation processing and stronger distractor inhibition processing than controls.[65] Keehn et al. (2008) used an event-related functional magnetic resonance imaging design to study the neurofunctional correlates of visual search in autistic children and matched controls of typically developing children.[69] Autistic children showed superior search efficiency and increased neural activation patterns in the frontal, parietal, and occipital lobes when compared to the typically developing children. Thus, autistic individuals’ superior performance on visual search tasks may be due to enhanced discrimination of items on the display, which is associated with occipital activity, and increased top-down shifts of visual attention, which is associated with the frontal and parietal areas.
Consumer psychology and visual search
In the past decade, there has been extensive research into how companies can maximise sales using psychological techniques, derived from visual search, to determine how products should be positioned on shelves. Pieters and Warlop (1999) [70] use eye tracking devices to assess saccades and fixations of consumers while they visually scanned/searched an array of products on a supermarket shelf. Their research suggests that consumers specifically direct their attention to products with eye-catching properties such as shape, colour or brand name. This is due to a pressured visual search where eye movements accelerate and saccades minimise, thus resulting in the consumer quickly choosing a product with a ‘pop out’ effect. This suggests that efficient search is primarily used concluding that consumers do not focus on items that share very similar features. Thus the more distinct or maximally visually different a product is from surrounding products, the more likely it is to be purchased by the consumer. Janiszewski (1998)[71] discussed two types of consumer search. One search type is goal directed search taking place when somebody uses stored knowledge of the product in order to make a purchase choice. The second is exploratory search. This occurs when the consumer has minimal previous knowledge about how to choose a product. It was found that for exploratory search, individuals would pay less attention to products that were placed in visually competitive areas such as the middle of the shelf at an optimal viewing height. This was primarily due to the competition in attention meaning that less information was maintained in visual working memory for these products.
References
- ↑ Shelga, B. M.; Riggio, L. & Rizzolatti, G. (1994). "Orienting of attention and eye movements". Experimental Brain Research 98: 507–522. doi:10.1007/bf00233988.
- ↑ Hoffman, J. E.; B. Subramaniam (1995). "The role of visual attention in saccadic eye movements". Perception and Psychophysics 57 (6): 787–795. doi:10.3758/bf03206794.
- ↑ 3.0 3.1 Treisman, A. M.; Gelade, G (1980). "A feature-integration theory of attention". . Cognitive psychology 12: 97–136. doi:10.1016/0010-0285(80)90005-5. PMID 7351125.
- ↑ Duncan, J; Humphreys, G, W. (1989). "Visual search and stimulus similarity". . Psychological Review 96 (3): 433–458. doi:10.1037/0033-295x.96.3.433.
- ↑ 5.0 5.1 5.2 Trick, L. M.; Enns, J. T. (1998). "Life-span changes in attention: The visual search task". Cognitive Development 13 (3): 369–386. doi:10.1016/s0885-2014(98)90016-8.
- ↑ Fernandez-Duque, D.; M. I. Posner (2001). "Brain imaging of attentional networks in normal and pathological states". Journal of Clinical and Experimental Neuropsychology 23 (1): 74–93. doi:10.1076/jcen.23.1.74.1217. Retrieved 2012-11-17.
- ↑ 7.0 7.1 Wurtz, Robert H.; Michael E. Goldberg; David Lee Robinson (June 1982). "Brain Mechanisms of Visual Attention". Scientific American 246 (6): 124–135. doi:10.1038/scientificamerican0682-124. ISSN 0036-8733. Retrieved 2012-11-17.
- ↑ 8.0 8.1 8.2 Müller, Hermann J.; Joseph Krummenacher (2006). "Visual search and selective attention". Visual Cognition 14 (4-8): 389–410. doi:10.1080/13506280500527676. ISSN 1350-6285. Retrieved 2012-11-09.
- ↑ Neisser, Ulric (1967). "Cognitive psychology.". Retrieved 2012-11-17.
- ↑ Treisman, A. M.; G. Gelade (1980). "A feature-integration theory of attention". Cognitive psychology 12 (1): 97–136. doi:10.1016/0010-0285(80)90005-5. PMID 7351125. Retrieved 2012-11-19.
- ↑ Wolfe, J. M. (1998). "What can 1 million trials tell us about visual search?". Psychological Science 9 (1): 33–39. doi:10.1111/1467-9280.00006. Retrieved 2012-11-20.
- ↑ 12.0 12.1 12.2 Wolfe, J. M. (1994). "Guided search 2.0 A revised model of visual search" (PDF). Psychonomic Bulletin & Review 1 (2): 202–238. doi:10.3758/bf03200774. Retrieved 2012-11-17.
- ↑ Aglioti, S.; Smania, N.; Barbieri, C.; Corbetta, M. (1997). "Influence of stimulus salience and attentional demands on visual search patterns in hemispatial neglect". Brain 34: 388–403. doi:10.1006/brcg.1997.0915.
- ↑ Eglin, M.; Robertson, L. C.; Knight, R. T. (1991). "Cortical substrates supporting visual search in humans.". Cerebral Cortex 1: 262–272. doi:10.1093/cercor/1.3.262.
- ↑ Friedman-Hill, S. R.,; Robertson, L. C.; Treisman, A. (1995). "Parietal contributions to visual feature binding: Evidence from a patient with bilateral lesions.". Science 269: 853–855. doi:10.1126/science.7638604.
- ↑ Ellison, A.,; Schindler, I.; Pattison, L. L.; Milner, A. D (2004). "An exploration of the role of the superior temporal gyrus in visualsearch and spatial perception using TMS.v". Brain, Advance Access.
- ↑ Ashbridge, V.,; Walsh, A. Cowey, D (1997). "Temporal aspects of visual search studied by transcranial magnetic stimulation". Neuropsychologia 35: 1121–1131. doi:10.1016/s0028-3932(97)00003-1.
- ↑ Nobre, A. C.; J. T. Coull; V. Walsh; C. D. Frith (2003). "Brain activations during visual search: contributions of search efficiency versus feature binding". Neuroimage 18 (1): 91–103. doi:10.1006/nimg.2002.1329.
- ↑ Leonards, U.; Suneart, S., Van Hecke, P., and Orban, G. (2000). "Attention mechanisms in visual search—An fMRI study". Journal of Cognitive Neuroscience 12: 61–75. doi:10.1162/089892900564073.
- ↑ Nobre, A.C,.; Sebestyen, G. N.; Gitelman, D. R.; Frith, C. D.; Mesulam, M. M. (2002). "Filtering of distractors during visual search studied by positron emission tomography". Neuroimage 16: 968–976. doi:10.1006/nimg.2002.1137.
- ↑ Schall JD. (2004). "On the role of frontal eye field in guiding attention and saccades". Vision Research 44 (12): 1453–1467. doi:10.1016/j.visres.2003.10.025. PMID 15066404.
- ↑ "Medical Neurosciences".
- ↑ Mustari MJ, Ono S, Das VE (May 2009). "Signal processing and distribution in cortical-brainstem pathways for smooth pursuit eye movements". Ann. N. Y. Acad. Sci. 1164: 147–54. doi:10.1111/j.1749-6632.2009.03859.x. PMC 3057571. PMID 19645893.
- ↑ McPeek, R.M,.; Keller, E. L., (2002). "Saccade target selection in the superior colliculus during a visual search task.". Journal of Neurophysiology 18: 2019–2034.
- ↑ Bender, D.B,.; Butter, C. M. (1987). "Comparison of the effects of superior colliculus and pulvinar lesions on visual search and tachistoscopic pattern discrimination in monkeys". Experimental Brain Research. 69: 140–154. doi:10.1007/bf00247037.
- ↑ Mazer, James A; Jack L Gallant (2003-12-18). "Goal-Related Activity in V4 during Free Viewing Visual Search: Evidence for a Ventral Stream Visual Salience Map". Neuron 40 (6): 1241–1250. doi:10.1016/S0896-6273(03)00764-5. ISSN 0896-6273. Retrieved 2012-11-20.
- ↑ Tomonaga, Masaki (2007-01-01). "Visual search for orientation of faces by a chimpanzee (Pan troglodytes): face-specific upright superiority and the role of facial configural properties". Primates 48 (1): 1–12. doi:10.1007/s10329-006-0011-4. ISSN 0032-8332. Retrieved 2012-11-20.
- ↑ LoBue, Vanessa; Judy S. DeLoache (2008-03-01). "Detecting the Snake in the Grass Attention to Fear-Relevant Stimuli by Adults and Young Children". Psychological Science 19 (3): 284–289. doi:10.1111/j.1467-9280.2008.02081.x. ISSN 0956-7976. Retrieved 2012-11-20.
- ↑ Goldberg, J. H.; C. K. Probart; R. E. Zak (1999). "Visual search of food nutrition labels". Human Factors: The Journal of the Human Factors and Ergonomics Society 41 (3): 425–437. doi:10.1518/001872099779611021. Retrieved 2012-11-20.
- ↑ Chapman, P. R.; G. Underwood (1998). "Visual search of driving situations: Danger and experience". Perception-London 27 (8): 951–964. doi:10.1068/p270951. Retrieved 2012-11-20.
- ↑ Janiszewski, C. (1998). "The influence of display characteristics on visual exploratory search behavior". Journal of Consumer Research 25 (3): 290–301. doi:10.1086/209540. Retrieved 2012-11-20.
- ↑ Kanwisher, Nancy; McDermott, Josh; Chun, Marvin M. "The fusiform face area: a module in human extrastriate cortex specialized for face perception". The Journal of Neuroscience 17 (11): 4302–4311.
- ↑ Tarr, M. J.; Gauthier, I. (2000). "FFA: a flexible fusiform area for subordinate-level visual processing automatized by expertise". Nature Neuroscience 3: 764–770.
- ↑ Grill-Spector, K.; Knouf, N.; Kanwisher, N. (2004). "The fusiform face area subserves face perception, not generic within-category identification". Nature Neuroscience 7 (5): 555–562. doi:10.1038/nn1224.
- ↑ Valentine, T; Bruce, V (1986). "The effects of distinctiveness in recognizing and classifying faces. Perception". Vol 15: 525–533. doi:10.1068/p150525.
- ↑ Purcell, D G; Stewart, A L (1986). "The face-detection effect. Bulletin of the Psychonomic Society". Vol 24: 118–120. doi:10.3758/bf03330521.
- ↑ Purcell, D G; Stewart, A L (1988). "The face-detection effect: Configuration enhances perception. Perception & Psychophysics". Vol 43: 355–366. doi:10.3758/bf03208806.
- ↑ Yovel, G.; Kanwisher, N. (2005). "The neural basis of the behavioural face-inversion effect". Current Biology 15: 2256–2262. doi:10.1016/j.cub.2005.10.072.
- ↑ Purcell, D G; Stewart, A L (1991). "The object-detection effect: Configuration enhances perception. Perception & Psychophysics". Vol 50: 215–224. doi:10.3758/bf03206744.
- ↑ Nothdurft, H. C. (1993). "Faces and facial expressions do not pop out". Perception 22 (11): 1287–98. doi:10.1068/p221287.
- ↑ Kuehn, S. M.; Jolicoeur, P. (1994). "Impact of quality of the image, orientation, and similarity of the stimuli on visual search for faces". Perception 23 (1): 95–122. doi:10.1068/p230095.
- ↑ Brown, V.; Huey, D.; Findlay, J. M. (1997). "Face detection in peripheral vision: do faces pop out?". Perception 26: 1555–1570. doi:10.1068/p261555.
- ↑ Nelson, C. A., (2001). The development and neural bases of face recognition. Infant and child development, 10 (1-2), 3-18. doi:10.1002/icd.239
- ↑ Hershler, O.; Hochstein, S. (2005). "At first sight: A high-level pop out effect for faces". Vision research 45 (13): 1707–1724. doi:10.1016/j.visres.2004.12.021.
- ↑ Hershler, O.; Golan, T.; Bentin, S.; Hochstein, S. (2010). "The wide window of face detection". Journal of vision 10 (10): 21. doi:10.1167/10.10.21.
- ↑ Simpson, E. A., Husband, H. L., Yee, K., Fullerton, A., & Jakobsen, K. V. (2014). Visual Search Efficiency Is Greater for Human Faces Compared to Animal Faces.
- ↑ VanRullen, R (2006). "On second glance: Still no high-level pop-out effect for faces". Vision research 46 (18): 3017–3027. doi:10.1016/j.visres.2005.07.009.
- ↑ Hershler, O.; Hochstein, S. (2006). "With a careful look: Still no low-level confound to face pop-out". Vision research 46 (18): 3028–3035. doi:10.1016/j.visres.2006.03.023.
- ↑ Golan, T.; Bentin, S.; DeGutis, J. M.; Robertson, L. C.; Harel, A. (2014). "Association and dissociation between detection and discrimination of objects of expertise: evidence from visual search". Attention, Perception, & Psychophysics 76 (2): 391–406. doi:10.3758/s13414-013-0562-6.
- ↑ Plude, D. J.; J. A. Doussard-Roosevelt (1989). "Aging, selective attention, and feature integration.". Psychology and Aging; Psychology and Aging 4 (1): 98–105. doi:10.1037/0882-7974.4.1.98. PMID 2803617. Retrieved 2012-11-19.
- ↑ Akhtar, N. (1990). "Peripheral vision in young children: Implications for the study of visual attention". The development of attention: Research and theory: 245–262. Retrieved 2012-11-19.
- ↑ Miller, L. K. (1973). "Developmental differences in the field of view during covert and overt search". Child Development: 247–252. Retrieved 2012-11-19.
- ↑ Enns, J. T.; D. A. Brodeur (1989). "A developmental study of covert orienting to peripheral visual cues". Journal of Experimental Child Psychology 48 (2): 171–189. doi:10.1016/0022-0965(89)90001-5. Retrieved 2012-11-19.
- ↑ Day, M. C. (1978). "Visual search by children: The effect of background variation and the use of visual cues". Journal of experimental child psychology 25 (1): 1–16. doi:10.1016/0022-0965(78)90034-6. Retrieved 2012-11-19.
- ↑ Harpur, L. L.; C. T. Scialfa; D. M. Thomas (1995). "Age differences in feature search as a function of exposure duration". Experimental Aging Research 21 (1): 1–15. doi:10.1080/03610739508254264. Retrieved 2012-11-19.
- ↑ Hartley, A. A.; J. M. Kieley; E. H. Slabach (1990). "Age differences and similarities in the effects of cues and prompts.". Journal of Experimental Psychology: Human Perception and Performance 16 (3): 523–537. doi:10.1037/0096-1523.16.3.523. Retrieved 2012-11-19.
- ↑ Connelly, S. L.; L. Hasher (1993). "Aging and the inhibition of spatial location.". Journal of Experimental Psychology: Human Perception and Performance 19 (6): 1238–1250. doi:10.1037/0096-1523.19.6.1238. Retrieved 2012-11-19.
- ↑ Rabbitt, P. (1965). "An age-decrement in the ability to ignore irrelevant information". Journal of Gerontology 20 (2): 233–238. doi:10.1093/geronj/20.2.233. Retrieved 2012-11-19.
- ↑ Lorenzo-López, L.; E. Amenedo; R. D. Pascual-Marqui; F. Cadaveira (2008). "Neural correlates of age-related visual search decline: a combined ERP and sLORETA study". NeuroImage 41 (2): 511–524. doi:10.1016/j.neuroimage.2008.02.041. Retrieved 2012-11-19.
- ↑ 60.0 60.1 60.2 60.3 Tales, A.; S. R. Butler; J. Fossey; I. D. Gilchrist; R. W. Jones; T. Troscianko (2002). "Visual search in Alzheimer’s disease: a deficiency in processing conjunctions of features". Neuropsychologia 40 (12): 1849–1857. doi:10.1016/S0028-3932(02)00073-8. Retrieved 2012-11-19.
- ↑ Parasuraman, R.; P. M. Greenwood; G. E. Alexander (2000). "Alzheimer disease constricts the dynamic range of spatial attention in visual search" (PDF). Neuropsychologia 38 (8): 1126–1135. doi:10.1016/s0028-3932(00)00024-5. Retrieved 2012-11-19.
- ↑ Mendez, M. F.; M. M. Cherrier; J. S. Cymerman (1997). "Hemispatial neglect on visual search tasks in Alzheimer's disease.". Neuropsychiatry, Neuropsychology, & Behavioral Neurology. Retrieved 2012-11-19.
- ↑ Troscianko, T.; J. Calvert (1993). "Impaired parallel visual search mechanisms in Parkinson's disease: implications for the role of dopamine in visual attention". Clinical vision sciences 8 (3): 281–287.
- ↑ Weinstein, A.; T. Troscianko; J. Calvert (1997). "Impaired visual search mechanisms in Parkinson's disease (PD): a psychophysical and event-related potentials study". Journal of Psychophysiology 11: 33–47.
- ↑ 65.0 65.1 O'Riordan, Michelle A.; Kate C. Plaisted; Jon Driver; Simon Baron-Cohen (2001). "Superior visual search in autism". Journal of Experimental Psychology: Human Perception and Performance 27 (3): 719–730. doi:10.1037/0096-1523.27.3.719. ISSN 1939-1277.
- ↑ 66.0 66.1 Remington, Anna M; John G Swettenham; Nilli Lavie (May 2012). "Lightening the load: perceptual load impairs visual detection in typical adults but not in autism". Journal of Abnormal Psychology 121 (2): 544–551. doi:10.1037/a0027670. ISSN 1939-1846.
- ↑ Remington, Anna; John Swettenham; Ruth Campbell; Mike Coleman (2009-11-01). "Selective Attention and Perceptual Load in Autism Spectrum Disorder". Psychological Science 20 (11): 1388–1393. doi:10.1111/j.1467-9280.2009.02454.x. ISSN 0956-7976. Retrieved 2012-12-20.
- ↑ Plaisted, Kate; Michelle O'Riordan; Simon Baron-Cohen (1998). "Enhanced Visual Search for a Conjunctive Target in Autism: A Research Note". Journal of Child Psychology and Psychiatry 39 (5): 777–783. doi:10.1111/1469-7610.00376. ISSN 1469-7610. Retrieved 2012-11-19.
- ↑ Keehn, Brandon; Laurie Brenner, Erica Palmer, Alan J. Lincoln, Ralph-Axel Müller (2008). "Functional brain organization for visual search in ASD". Journal of the International Neuropsychological Society 14 (06): 990–1003. doi:10.1017/S1355617708081356.
- ↑ Pieters, R.,; Warlop, L. (1999). "Visual attention during brand choice: the impact of time pressure and task motivation.". International journal of research in marketing. Elsevier Science. 16: 1–16. doi:10.1016/s0167-8116(98)00022-6.
- ↑ Janiszewski, C., (1998). "The Influence of Display Characteristics on Visual Exploratory Search Behavior.". Journal of Consumer Research. 25 (3): 290–301. doi:10.1086/209540.
External links
- http://www.gardnergallery.com/ Some fun visual search tasks by artist Steven Gardner.
- Google Goggles An artificial visual search application.
- Picalike Specialized company on visual search technology
- LikeThat One of the first implementations for visual-search in shopping