Traumatic insemination

Traumatic insemination, also known as hypodermic insemination, is the mating practice in some species of invertebrates in which the male pierces the female's abdomen with his penis and injects his sperm through the wound into her abdominal cavity (hemocoel).[1] The sperm diffuse through the female's hemolymph, reaching the ovaries and resulting in fertilization. The process is detrimental to the female's health. It creates an open wound which impairs the female until it heals, and is susceptible to infection. The injection of sperm and ejaculatory fluids into the hemocoel can also trigger an immune reaction in the female. Bed bugs, which reproduce solely by traumatic insemination, have evolved a pair of sperm-receptacles, known as the spermalege. It has been suggested that the spermalege reduces the direct damage to the female bed bug during traumatic insemination. However, statistical experiments showed no conclusive evidence for that hypothesis; hygienic protection against bacteria being the preferred explanation for that organ.[2]
The evolutionary origins of traumatic insemination are disputed. Although it evolved independently in many invertebrate species, traumatic insemination is most highly adapted and thoroughly studied in bed bugs, particularly Cimex lectularius.[1][3] Traumatic insemination is not limited to male-female couplings, or even couplings of the same species. Both homosexual and inter-species traumatic inseminations have been observed.
Mechanics

In humans and other complex life forms, blood and lymph circulate in two different systems, the circulatory system and lymphatic system, which are enclosed by systems of capillaries, veins, arteries, and nodes. This is known as a closed circulatory system. Insects, however, have an open circulatory system in which blood and lymph circulate unenclosed, and mix to form a substance called hemolymph. All organs of the insect are bathed in hemolymph, which provides oxygen and nutrients to all of the insect's organs.[4]
Following traumatic insemination, sperm can migrate through the hemolymph to the female's ovaries, resulting in fertilization. The exact mechanics vary from taxon to taxon. In some orders of insects, the male genitalia (paramere) enters the female's genital tract, and a spine at its tip pierces the wall of the female's bursa copulatrix. In others, the male penetrates the outer body wall. In either case, following penetration, the male ejaculates into the female. The sperm and ejaculatory fluids diffuse through the female's hemolymph. The insemination is successful if the sperm reach the ovaries and fertilize an ovum.[1]
Female resistance to traumatic insemination varies from one species to another. Females from some genera, including Cimex, are passive prior to and during traumatic insemination.[5] Females in other genera resist mating and attempt to escape.[6] This resistance may not be an aversion to pain caused by the insemination, as observational evidence suggests that insects do not feel pain.[7]
Research into the paternity of offspring produced by traumatic insemination has found "significant" last-sperm precedence. That is, the last male to traumatically inseminate a female tends to sire most of the offspring from that female.[8]
Evolutionary adaptation
Many reasons for the evolutionary adaptation of traumatic insemination as a mating strategy have been suggested. One is that traumatic insemination is an adaptation to the development of the mating plug, a reproductive mechanism used by many species. Once a male finishes copulating, he injects a glutinous secretion into the female's reproductive tract, thereby "literally glu[ing] her genital tract closed".[9] Traumatic insemination allows subsequent males to bypass the female's plugged genital tract, and inject sperm directly into her circulatory system.
Others have argued that the practice of traumatic insemination may have been an adaptation for males to circumvent female resistance to mating[1] to eliminate courtship time, allowing one male to inseminate many mates when contact between them is brief;[10] or that it evolved as a new development in the sperm competition as a means to deposit sperm as close to the ovaries as possible.
This bizarre method of insemination probably evolved as male bed bugs competed with each other to place their sperm closer and closer to the mother lode of eggs, the ovaries. Some male insects evolved long penises with which they enter the vagina but bypass the female's storage pouch and deposit their sperm further upstream close to the ovaries. A few males, notably among bed bugs, evolved traumatic insemination instead, and eventually this strange procedure became the norm among these insects.[11]
It has recently been discovered that members of the plant bug genus Coridromius (Miridae) also practice traumatic insemination. In these bugs, the male intromittent organ is formed by the coupling of the aedeagus with the left paramere, as in bed bugs. Females also exhibit paragenital modifications at the site of intromission, which include grooves and invaginated copulatory tubes to guide the male paramere. The evolution of traumatic insemination in Coridromius represents a third independent emergence of this form of mating within the true bugs.[12]
Health repercussions
While advantageous to the reproductive success of the individual male, traumatic insemination imposes a cost on females: reduced lifespan and decreased reproductive output. "These [costs] include (i) repair of the wound, (ii) leakage of blood, (iii) increased risk of infection through the puncture wound, and (iv) immune defence against sperm or accessory gland fluids that are introduced directly into the blood."[1]
The male bed bug penis has been shown to carry five (human) pathogenic microbes, and the exoskeleton of female bed bugs nine, including Penicillium chrysogenum, Staphylococcus saprophyticus, Stenotrophomonas maltophilia, Bacillus licheniformis, and Micrococcus luteus. Tests with blood agar have shown some of these species can survive in vivo. This suggests infections from these species may contribute to the increased mortality rate in bed bugs due to traumatic insemination.[13]
The successive woundings each require energy to heal, leaving less energy available for other activities. Also, the wounds provide a possible point of infection which can reduce the female's lifespan.[8] Once in the hemolymph, the sperm and ejaculatory fluids may act as antigens, triggering an immune reaction.
There is a tendency for dense colonies of bed bugs kept in laboratories to go extinct, starting with adult females.[14] In such an environment, where mating occurs frequently, this high rate of adult female mortality suggests traumatic insemination is very detrimental to the female's health.[1] The damage done, and the (unnecessarily) high mating rate of captive bed bugs, have been shown to cause a 25% higher-than-necessary mortality rate for females.[15]
Bed bug adaptation

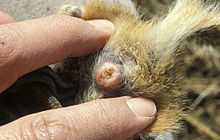
The effects of traumatic insemination are deleterious to the female. Female bed bugs have evolved a pair of specialized reproductive organs ("paragenitalia") at the site of penetration. Known as the ectospermalege and mesospermalege (referred to collectively as spermalege), these organs serve as sperm-receptacles from which sperm can migrate to the ovaries.[2][16] All bed bug reproduction occurs via traumatic insemination and the spermalege. The genital tract, though functional,[17] is used only for laying fertilized eggs.[3]
The ectospermalege is a swelling in the abdomen, often folded, filled with hemocytes. The ectospermalege is visible externally in most bed bug species, giving the male a target through which to impale the female with the paramere. In species without an externally visible ectospermalege, traumatic insemination takes place over a wide range of the body surface.[17]
Exactly why males 'comply' with this aspect of female control over the site of mating is unclear, especially as male P. cavernis appear to be able to penetrate the abdomen at a number of points independent of the presence of an ectospermalege. One possibility is that mating outside the ectospermalege reduces female fecundity to such an extent that the mating male's paternity is significantly reduced ... The ectospermalege appears to act as a mating guide, directing the male's copulatory interest, and therefore damage, to a restricted area of the female's abdomen.[17]
The mesospermalege is a sac attached to the inner abdomen, under the ectospermalege. Sperm is injected through the male's penis into the mesospermalege. In some species, the ectospermalege directly connects to the ovaries – thus, sperm and ejaculate never enters the hemolymph and thus never trigger an immune reaction. (The exact characteristics of the spermalege vary widely across different species of bed bugs.)[1] The spermalege are generally found only in females. However, males in the Afrocimex genus possess an ectospermalege.[1] Sperm remains in the spermalege for approximately four hours; after two days, none remains.[3]
Male bed bugs have evolved chemoreceptors on their penises. After impaling a female, the male can "taste" if a female has been recently mated. If he does, he will not copulate as long and will ejaculate less fluid into the female.[18]
Use in the animal kingdom
Although traumatic insemination is most widely practiced among heteropterans (typical bugs), the phenomenon has been observed across a wide variety of other invertebrate taxa. These include:
- Oxyurida (nematodes) – Traumatic insemination has been observed in pinworm genera including Auchenacantha, Citellina, Passalurus, and "probably" Austroxyris.[1][19][20]
- Acanthocephala (parasitic, thorny-headed worms) – The presence of mating plugs on the sides of Pomphorhynchus bulbocolli suggests traumatic insemination occurs in this species. Because these parasites cannot move after anchoring themselves to a host's intestine, traumatic insemination may have evolved to compensate for their immobility.[1][21]
- Rotifera (wheel animalcules) – In the Brachionus genus, the male pierces the syncytial integument (equivalent to skin) and injects sperm; in Asplanchna brightwelli the male secretes an enzyme which breaks down the female integument and injects sperm through the hole.[22][23]
- Turbellaria (free living flatworms) – Hermaphroditic flatworms reproduce by "penis fencing". Individuals "fence" with penises, attempting to use their penis to pierce the skin of the other and inject sperm. The 'loser' is the flatworm which is inseminated and must bear the energy costs of reproduction. One study of Pseudoceros bifurcus found "Most inseminations were unilateral. Even when reciprocal penis insertion could be achieved by the second partner, the first to inseminate obtained a longer injection time than the second."[24][25]
- Gastropod snails[1]
- Strepsiptera (twisted-winged parasites) – In Xenos vesparum, fertilization can occur either via extragenital ducts, or by traumatic insemination into the hemocoel.[26]
- Drosophila (fruit flies) – Ejaculates are injected through the body wall into the genital tract, not the abdomen.[27]
- Opisthobranchia (sea slugs) – Characterized by "repeated small injections into the dorsal surface of the partner, interrupted by synchronised circling movements", culminating in a standard genital insemination.[28]
- Harpactea (spiders) – The male of the spider species Harpactea sadistica pierces the female's body cavity and inseminates her ovaries directly.[29]
Homosexual traumatic insemination
Traumatic insemination is not limited to male–female couplings. Male homosexual traumatic inseminations have been observed in the flower bug Xylocoris maculipennis[9] and bed bugs of the genus Afrocimex.[1]
In the genus Afrocimex, both species have well developed ectospermalege (but only females have a mesospermalege). The male ectospermalege is slightly different from that found in females, and amazingly enough, Carayon (1966) found that male Afrocimex bugs suffer actual homosexual traumatic inseminations. He found the male ectospermalege often showed characteristic mating scars, and histological studies showed "foreign" sperm were widely dispersed in the bodies of these homosexually mated males. Sperm cells of other males were, however, never found in or near the male reproductive tract. It therefore seems unlikely that sperm from other males could be inseminated when a male that has himself suffered traumatic insemination mates with a females. The costs and benefits, if any, of homosexual traumatic insemination in Afrocimex remain unknown.[1]
Klaus Reinhardt of the University of Sheffield and colleagues observed two morphologically different kinds of spermalege in Afrocimex constrictus, a species in which both male and females are traumatically inseminated. They found females use sexual mimicry as a way to avoid traumatic insemination. In particular, they observed males, and females who had male spermalege structures, were inseminated less often than females with female spermalege structures.[30]
In Xylocoris maculipennis, after a male traumatically inseminates another male, the injected sperm migrate to the testes. (The seminal fluid and most of the sperm are digested, giving the inseminated male a nutrient-rich meal.) It has been suggested, although there is no evidence, that when the inseminated male ejaculates into a female, the female receives both males' sperm.[31][32]
Interspecies traumatic insemination
Cases of traumatic insemination between animals of different species will sometimes provoke a possibly lethal immune reaction. A female Cimex lectularius traumatically inseminated by a male C. hemipterus will swell up at the site of insemination as the immune system responds to male ejaculates. In the process, the female's lifespan is reduced. In some cases, this immune reaction can be so massive as to be almost immediately fatal. A female Hesperocimex sonorensis will swell up, blacken, and die within 24–48 hours after being traumatically inseminated by a male H. cochimiensis.[1]
Similar mating practices
In the animal kingdom, traumatic insemination is not unique as a form of coercive sex. Research suggests, in the Acilius genus of water beetles, there is no courtship system between males and females. "It's a system of rape. But the females don't take things quietly. They evolve counter-weapons." Cited mating behaviors include males suffocating females underwater till exhausted, and allowing only occasional access to the surface to breathe for up to six hours (to prevent them breeding with other males), and females which have a variety of body shapes (to prevent males from gaining a grip). Foreplay is "limited to the female desperately trying to dislodge the male by swimming frantically around".[33]
"Rape behavior" has been observed in a number of duck species. In the blue-winged teal, "rape attempts by paired males may occur at any time during the breeding season." Cited reasons for this being beneficial to the paired males include successful reproduction, and chasing away intruders from their territory.[34] Bachelor herds of bottlenose dolphins will sometimes gang up on a female and coerce her to have sex with them, by swimming near her, chasing her if she attempts to escape, and making vocalized or physical threats.[35][36] In the insect world, male water striders unable to penetrate her genital shield, will draw predators to a female until she copulates.[37]
See also
References
- ↑ 1.0 1.1 1.2 1.3 1.4 1.5 1.6 1.7 1.8 1.9 1.10 1.11 1.12 1.13 Arnqvist, Göran; Rowe, Locke (2005-07-05). Sexual Conflict (Monographs in Behavior and Ecology). Princeton, NJ: Princeton University Press. pp. 87–91. ISBN 978-0-691-12218-2.
- ↑ 2.0 2.1 Reinhardt, K.; Naylor R.; Siva-Jothy, M. T. (2003-11-22). "Reducing a cost of traumatic insemination: female bedbugs evolve a unique organ". Proceedings of the Royal Society B 270 (1531): 2371–2375. doi:10.1098/rspb.2003.2515. PMC 1691512. PMID 14667353.
- ↑ 3.0 3.1 3.2 Carayon, J. (1966). "Paragenital system". Monograph of Cimicidae (Hemiptera, Heteroptera). College Park, MD: Entomological Society of America. pp. 81–166. ASIN B0000EGQ6L. ISBN 978-0-9776209-2-0.
- ↑ Ruck, B.N.; Villares, G.J.; Welch, C.M.; Facey, D.E. "Circulatory System". Saint Michael's College. Retrieved 2009-08-19.
- ↑ Arnqvist, citing A.D. Stutt, Reproductive Strategies and Sexual Conflict in the Bed Bug. PhD thesis. University of Sheffield, England.
- ↑ Arnqvist 89, citing personal observations and Monograph of Cimicidae
- ↑ Smith, Jane A. (1991). "A Question of Pain in Invertebrates". ILAR Journal 33 (1–2). Retrieved 2009-04-07.
In the majority of examples of invertebrate nociception noted above, there seems to be little, if any, evidence that the animals' responses persist in anything akin to the manner described for mammals. As Eisemann et al. (1984) have described in a review of the 'biological evidence' concerning pain in insects, 'No example is known to us of an insect showing protective behavior towards injured parts, such as by limping after leg injury or declining to feed or mate because of general abdominal injuries. On the contrary, our experience has been that insects will continue with normal activities even after severe injury or removal of body parts.'... Certainly, on the limited amount of evidence presented here, it seems very difficult to imagine that insects and the other simpler invertebrates mentioned above can 'suffer' pain in anything like the vertebrate sense. Nevertheless, the issue certainly is not closed, and further questions should be asked.
- ↑ 8.0 8.1 Stutt AD, Siva-Jothy MT (2001-05-08). "Traumatic insemination and sexual conflict in the bed bug Cimex lectularius". Proceedings of the National Academy of Sciences, U.S.A. 98 (10): 5683–87. doi:10.1073/pnas.101440698. PMC 33273. PMID 11331783. Retrieved 2009-04-09.
- ↑ 9.0 9.1 Quammen, David (1998-02-16) [1988]. The Flight of the Iguana: A Sidelong View of Science and Nature. Simon & Schuster. pp. 29–30. ISBN 978-0-684-83626-3.
- ↑ Trowbridge, C. D. (1995). "Hypodermic insemination, oviposition, and embryonic development of a pool-dwelling ascoglossan (=sacoglossan) opisthobranch: Ercolania felina (Hutton, 1882) on New Zealand shores". The Veliger 38: 203–211.
- ↑ Waldbauer, Gilbert (1998-02-11). Insects through the seasons. Harvard University Press. p. 63. ISBN 978-0-674-45489-7.
- ↑ Tatarnic, Nikolai J; Gerasimos Cassis; Dieter F Hochuli (2006-03-22). "Traumatic insemination in the plant bug genus Coridromius Signoret (Heteroptera: Miridae)". Biol Lett. (The Royal Society) 2 (1): 58–61. doi:10.1098/rsbl.2005.0394. PMC 1617170. PMID 17148326.
- ↑ Reinhardt, Klaus; Naylor, Richard A.; Siva-Jothy, Michael T. (2005-09-21). "Potential sexual transmission of environmental microbes in a traumatically inseminating insect". Ecological Entomology 30 (5): 607–611. doi:10.1111/j.0307-6946.2005.00730.x.
- ↑ Arnqvist, citing Monograph of Cimicidae
- ↑ "University of Sheffield Department of Animal & Plant Sciences Research Projects". Retrieved 2004-09-19.
For this purpose we are studying the traumatic insemination of bed bugs. The natural mating rate of bed bugs is such that females suffer 25% higher mortality than is necessary for full fertilization. This is apparently not outweighed by 25% benefits to the offspring. So far, we have identified that a unique organ, the spermalege has possibly evolved to mediate pathogenic microbes transmitted during copulation. By contrast, the amount of wounding and wound repair in response to piercing is not mediated by the spermalege, nor are the action of seminal fluids. We are presently testing whether costs to activate the immune system, or costs to maintain sperm alive contribute to the increase female mortality.
- ↑ Morrow, Edward H.; Arnqvist, Göran (2003-11-22). "Costly traumatic insemination and a female counter-adaptation in bed bugs". Proceedings of the Royal Society B 270 (1531): 2377–2381. doi:10.1098/rspb.2003.2514. PMC 1691516. PMID 14667354.
- ↑ 17.0 17.1 17.2 Siva-Jothy MT (2006-02-28). "Trauma, disease and collateral damage: conflict in cimicids". Philosophical Transactions of the Royal Society B 361 (1466): 269–75. doi:10.1098/rstb.2005.1789. PMC 1569606. PMID 16612886.
- ↑ Siva-Jothy MT and Alistair D. Stutt (March 2003). "A matter of taste: direct detection of female mating status in the bedbug". Proceedings of the Royal Society B 270 (1515): 649–652. doi:10.1098/rspb.2002.2260. PMC 1691276. PMID 12769466.
- ↑ Anderson, Roy Clayton (2000). Nematode parasites of vertebrates. CABI. p. 233. ISBN 0-85199-421-0.
- ↑ Hugot, JP (1982-04-19). "Traumatic insemination and tubular egg receptacle in the pinworm [Passalurus ambigus] parasite of the common domestic rabbit". Comptes Rendus Hebdomadaires des Seances de l'Academie des Sciences. Serie 3. Sciences de la Vie (France) 294 (14): 707–10.
- ↑ Doyle, Lester R.; Larry N. Gleason (June 1991). "Suckers and Other Bursal Structures of Pomphorhynchus bulbocolli and Acanthocephalus dirus (Acanthocephala)". The Journal of Parasitology (The American Society of Parasitologists) 77 (3): 437–40. doi:10.2307/3283132. JSTOR 3283132. PMID 2040955.
- ↑ Wallace, R. L. (1998). "Rotifera". In Knobil, Ernst; Neill, Jimmy D. Encyclopedia of Reproduction. Boston: Academic Press. pp. 290–301. ISBN 0-12-227020-7.
- ↑ Obst, Matthias; Funch, Peter (2003-01-06). "Dwarf male of symbion pandora (cycliophora)". Journal of Morphology 255 (3): 261–78. doi:10.1002/jmor.10040. PMID 12520546.
- ↑ Michiels, N. K.; Newman, L. J. (1998-02-12). "Sex and violence in hermaphrodites". Nature 391 (647): 647–8. doi:10.1038/35527. ISSN 0028-0836. PMID 859632.
- ↑ Milius, S. "Hermaphrodites duel for manhood". Science News Online. Retrieved 2009-08-19.
- ↑ Beani L, Giusti F, Mercati D, Lupetti P, Paccagnini E, Turillazzi S, Dallai R (2005-07-26). "Mating of Xenos vesparum (Rossi) (Strepsiptera, Insecta) revisited". Journal of Morphology 265 (3): 291–303. doi:10.1002/jmor.10359. PMID 16047336.
- ↑ Kamimura, Yoshitaka (2007-08-22). "Twin intromittent organs of Drosophila for traumatic insemination". Biol Lett. (The Royal Society) 3 (4): 401–404. doi:10.1098/rsbl.2007.0192. PMC 2391172. PMID 17519186.
- ↑ Schmitt V, Anthes N, Michiels NK (2007). "Mating behaviour in the sea slug Elysia timida (Opisthobranchia, Sacoglossa): hypodermic injection, sperm transfer and balanced reciprocity". Frontiers in Zoology 4: 17. doi:10.1186/1742-9994-4-17. PMC 1934903. PMID 17610714. Retrieved 2009-04-09.
- ↑ Řezáč, Milan (2009-04-29). "The spider Harpactea sadistica: co-evolution of traumatic insemination and complex female genital morphology in spiders". Proceedings of the Royal Society B 276 (1668): 2697–701. doi:10.1098/rspb.2009.0104. PMC 2839943. PMID 19403531. Retrieved 2009-05-09. Abstract at link
- ↑ Reinhardt, Klaus; Ewan Harney, Richard Naylor, Stanislav Gorb, Michael T. Siva‐Jothy (2007). "Female‐Limited Polymorphism in the Copulatory Organ of a Traumatically Inseminating Insect". American Naturalist 170 (6): 931–35. doi:10.1086/522844. PMID 18171174.
- ↑ Krebs, John R.; Davies, Nicholas B. (1993). An Introduction to Behavioural Ecology. Wiley-Blackwell. p. 186. ISBN 978-0-632-03546-5., citing Carayon, J. (1974). "Insemination traumatique heterosexuelle et homosexuelle chez Xylocoris maculipennis (Hem., Anthocoridae)". Comptes Rendus de l'Academie des Sciences (France) 278: 2803–06.
- ↑ Abele, Lawrence G.; Sandra Gilchrist (1977). "Homosexual Rape and Sexual Selection In Acanthocephalan Worms". Science 197 (4298): 81–83. doi:10.1126/science.867055. PMID 867055.
- ↑ Smith, Lewis (2007-06-25). "Not tonight, not ever. I've got a headache. Don't come near me". The Times (London). p. 25. Retrieved 2009-04-03.
- ↑ Bailey, Robert O.; Seymour, Norman R.; Stewart, Gary R. (January 1978). "Rape Behavior in Blue-Winged Teal". The Auk 95 (1): 188–90. doi:10.2307/4085514.
- ↑ Angier, Natalie (1992-02-18). "Dolphin Courtship: Brutal, Cunning and Complex". The New York Times (New York City). p. C1. Retrieved 2009-04-03.
- ↑ Connor, Richard C.; F. Andrew; Rachel A. Smolker; Janet Mann (February 1996). "Patterns of Female Attractiveness in Indian Ocean Bottlenose Dolphins". Behaviour 133 (1/2): 37–69. doi:10.1163/156853996X00026.
- ↑ Han, Chang S.; Jablonski, Piotr G. (August 2010). "Male water striders attract predators to intimidate females into copulation". Nature Communications 1 (52): 1. doi:10.1038/ncomms1051. PMC 2964456. PMID 20975717.