Transmission disequilibrium test
The transmission disequilibrium test (TDT) was proposed by Spielman, McGinnis and Ewens (1993)[1] as a family-based association test for the presence of genetic linkage between a genetic marker and a trait. It is an application of McNemar's test.
A specificity of the TDT is that it will detect genetic linkage only in the presence of genetic association. While genetic association can be caused by population structure, genetic linkage will not be affected, which makes the TDT robust to the presence of population structure.
The case of trios: one affected child per family
Description of the test
We first describe the TDT in the case where families consist of trios (two parents and one affected child). Our description follows the notations used in Spielman, McGinnis & Ewens (1993).[1]
The TDT measures the over-transmission of an allele from heterozygous parents to affected offsprings.
The n affected offsprings have 2n parents. These can be represented by the transmitted and the non-transmitted alleles 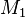 and
and 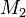 at some genetic locus. Summarizing the data in a 2 by 2 table gives:
at some genetic locus. Summarizing the data in a 2 by 2 table gives:
| Non-transmitted allele | |||
|---|---|---|---|
| Transmitted allele | M1 | M2 | Total |
| M1 | a | b | a + b |
| M2 | c | d | c + d |
| Total | a + c | b + d | 2n |
The derivation of the TDT shows that one should only use the heterozygous parents (total number b+c). The TDT tests whether the proportions b/(b+c) and c/(b+c) are compatible with probabilities (0.5, 0.5). This hypothesis can be tested using a binomial (asymptotically chi-square) test with one degree of freedom:
![\chi^2 = \frac{ [b - (b+c)/2]^2}{(b+c)/2} + \frac{ [c - (b+c)/2]^2}{(b+c)/2} = \frac{(b-c)^2}{b+c}](../I/m/560cfbc15855e9189f7f05798ba235d7.png)
Outline of the test derivation
A derivation of the test consists of using a population genetics model to obtain the expected proportions for the quantities 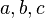 and
and 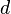 in the table above. In particular, one can show that under nearly all disease models the expected proportion of
in the table above. In particular, one can show that under nearly all disease models the expected proportion of 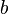 and
and  are identical. This result motivates the use of a binomial (asymptotically
are identical. This result motivates the use of a binomial (asymptotically 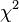 ) test to test whether these proportions are equal.
) test to test whether these proportions are equal.
On the other hand, one can also show that under such models the proportions and are not equal to the product of the marginals probabilities 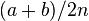 ,
, 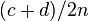 and
and 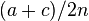 ,
, 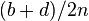 . A rewording of this statement would be that the type of the transmitted allele is not, in general, independent of the type of the non-transmitted allele. A consequence is that a test for homogeneity/independence does not test the appropriate hypothesis, and thus, only heterozygous parents are included.
. A rewording of this statement would be that the type of the transmitted allele is not, in general, independent of the type of the non-transmitted allele. A consequence is that a test for homogeneity/independence does not test the appropriate hypothesis, and thus, only heterozygous parents are included.
Extension to two affected child per family
Extension of the test
The TDT can be readily extended beyond the case of trios. We keep following the notations of Spielman, McGinnis & Ewens (1993).[1] Let us consider a total of 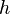 heterozygous parents. We use the fact that the transmission to different children are independent. The information can be then summarized in three categories:
heterozygous parents. We use the fact that the transmission to different children are independent. The information can be then summarized in three categories:
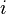 = number of parents who transmit to both children.
= number of parents who transmit to both children.
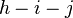 = number of parents who transmit to one child and to another.
= number of parents who transmit to one child and to another.
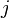 = number of parents who transmit to both children.
= number of parents who transmit to both children.
Using the notations of the previous paragraph we have:
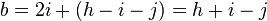
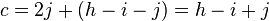
leading to the chi-squared test statistic:
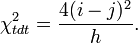
Relation with another linkage statistic
The comparison with the more traditional (at least at the time when the TDT was proposed) linkage test proposed by Blackwelder and Elston 1985[2] is informative. The Blackwelder and Elston approach uses the total number of haplotypes identical by descent (mean haplotype sharing). This measure ignores the allelic state of a marker and simply compares the number of times a parent transmits the same allele to both affected children with the number of times a different allele is transmitted. The test statistic is:
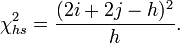
Under the null hypothesis of no linkage the expected proportions of (i, h − i − j, j) are (0.25, 0.5, 0.25). One can derive a simple chi-square statistic with 2 degrees of freedom:
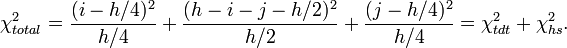
It clearly appears that the total statistic (with two degree of freedom) is the sum of two independent components: one is the traditional linkage measure and the other is the TDT statistic.
A modified version of the TDT
More recently, Wittkowski KM, Liu X. (2002/2004)[3] proposed a modification to the TDT that can be more powerful under some alternatives, although the asymptotic properties under the null hypothesis are equivalent.
The motivating idea for this modification is the fact that, while the transmissions of both allele from parents to a child are independent, the effects of other filial genetic or environmental covariates on penetrance are the same for both alleles transmitted to the same child. This situation can be important if, for example, the genetic marker is linked to a disease locus with a strong selection against heterozygous individuals. This observation suggests to shift the statistical model from a set of independent transmissions to a set of independent children (see Sasieni (1997)[4] for the corresponding problem in case-control association tests). While this observation does not affect the distribution under the null hypothesis of no linkage, it allows, for some disease models, to design a more powerful test.
In this modified TDT test the children are stratified by parental type and the modified test statistic becomes:
![\chi^2 = \frac{ \left[ [n_{PQ} - n_{QQ}]_{PQ \sim QQ} + 2\times[n_{PP} - n_{QQ}]_{PQ \sim PQ} + [n_{PP} - n_{PQ}]_{PP \sim PQ} \right]^2}{[n_{PQ} + n_{QQ}]_{PQ \sim QQ} + 4\times[n_{PP} + n_{QQ}]_{PQ \sim PQ} + [n_{PQ} + n_{PP}]_{PP \sim PQ}}](../I/m/a9bd254f1d426df99ba3cd1c9ccbb96b.png)
where ![[n_{PQ}]_{PQ \sim QQ}](../I/m/9f6a5bfd240b74f4c942935a64f4f585.png) is the number of PQ children from parents with the PQ and QQ types.
is the number of PQ children from parents with the PQ and QQ types.
References
- ↑ 1.0 1.1 1.2 Spielman RS, McGinnis RE, Ewens WJ (Mar 1993). "Transmission test for linkage disequilibrium: the insulin gene region and insulin-dependent diabetes mellitus (IDDM)". Am J Hum Genet. 52 (3): 506–16. PMC 1682161. PMID 8447318.
- ↑ Blackwelder WC, Elston RC (1985). "A comparison of sib-pair linkage tests for disease susceptibility loci". Genetic epidemiology 2 (1): 85–97. doi:10.1002/gepi.1370020109. PMID 3863778.
- ↑ Wittkowski KM, Liu X (2002). "A statistically valid alternative to the TDT". Hum Hered. 54 (3): 157–64. doi:10.1159/000068840. PMID 12626848.
Ewens WJ, Spielman RS (2004). "The TDT is a statistically valid test: comments on Wittkowski and Liu". Hum Hered. 58 (1): 59–60; author reply 60–1; discussion 61–2. doi:10.1159/000081458. PMID 15604566. - ↑ Sasieni PD (Dec 1997). "From genotypes to genes: doubling the sample size". Biometrics 53 (4): 1253–61. doi:10.2307/2533494. JSTOR 2533494. PMID 9423247.
- Ewens WJ, Spielman RS (2005). "What is the significance of a significant TDT?". Hum Hered. 60 (4): 206–10. doi:10.1159/000090544. PMID 16391488.
- Spielman RS, Ewens WJ (Feb 1998). "A sibship test for linkage in the presence of association: the sib transmission/disequilibrium test". Am J Hum Genet. 62 (2): 450–8. doi:10.1086/301714. PMC 1376890. PMID 9463321.
- Spielman RS, Ewens WJ (Nov 1996). "The TDT and other family-based tests for linkage disequilibrium and association". Am J Hum Genet 59 (5): 983–9. PMC 1914831. PMID 8900224.
- Ewens WJ, Spielman RS (Aug 1995). "The transmission/disequilibrium test: history, subdivision, and admixture". Am J Hum Genet. 57 (2): 455–64. PMC 1801556. PMID 7668272.
- McGinnis RE, Ewens WJ, Spielman RS (1995). "The TDT reveals linkage and linkage disequilibrium in a rare disease". Genet Epidemiol. 12 (6): 637–40. doi:10.1002/gepi.1370120619. PMID 8787986.