Transcription activator-like effector nuclease
Transcription activator-like effector nucleases (TALENs) are artificial restriction enzymes generated by fusing a TAL effector DNA-binding domain to a DNA cleavage domain.
Restriction enzymes are enzymes that cut DNA strands at a specific sequence. Transcription activator-like effectors (TALEs) can be quickly engineered to bind practically any desired DNA sequence.[1] By combining such an engineered TALE with a DNA cleavage domain (which cuts DNA strands), one can engineer restriction enzymes that are specific for any desired DNA sequence. When these restriction enzymes are introduced into cells, they can be used for genome editing in situ, a technique known as genome editing with engineered nucleases.
TALE DNA-binding domain
TAL effectors are proteins secreted by Xanthomonas bacteria. The DNA binding domain contains a repeated highly conserved 33–34 amino acid sequence with the exception of the 12th and 13th amino acids. These two locations are highly variable (Repeat Variable Diresidue) and show a strong correlation with specific nucleotide recognition.[2][3] This simple relationship between amino acid sequence and DNA recognition has allowed for the engineering of specific DNA-binding domains by selecting a combination of repeat segments containing the appropriate RVDs.[1]
DNA cleavage domain
The non-specific DNA cleavage domain from the end of the FokI endonuclease can be used to construct hybrid nucleases that are active in a yeast assay.[4][5] These reagents are also active in plant cells[6][7] and in animal cells.[7][8][9][10] Initial TALEN studies used the wild-type FokI cleavage domain, but some subsequent TALEN studies[9][11][12] also used FokI cleavage domain variants with mutations designed to improve cleavage specificity[13][14] and cleavage activity.[15] The FokI domain functions as a dimer, requiring two constructs with unique DNA binding domains for sites in the target genome with proper orientation and spacing. Both the number of amino acid residues between the TALE DNA binding domain and the FokI cleavage domain and the number of bases between the two individual TALEN binding sites appear to be important parameters for achieving high levels of activity.[8][16]
Engineering TALENs
The simple relationship between amino acid sequence and DNA recognition of the TALE binding domain allows for designable proteins. In this case artificial gene synthesis is problematic because of improper annealing of the repetitive sequence found in the TALE binding domain.[17] One solution to this is to use a publicly available software program (DNAWorks) to calculate oligonucleotides suitable for assembly in a two step PCR; oligonucleotide assembly followed by whole gene amplification. A number of modular assembly schemes for generating engineered TALE constructs have also been reported.[7][17][18][19][20][21] Both methods offer a systematic approach to engineering DNA binding domains that is conceptually similar to the modular assembly method for generating zinc finger DNA recognition domains.
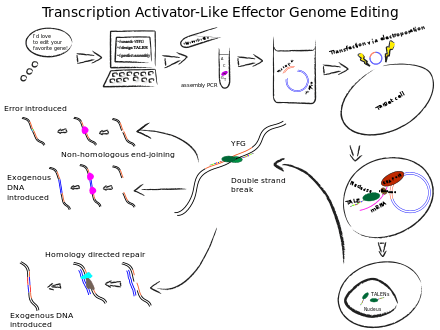
Transfection
Once the TALEN genes have been assembled they are inserted into plasmids; the plasmids are then used to transfect the target cell where the gene products are expressed and enter the nucleus to access the genome. Alternatively, TALENs can be delivered to the cell as mRNA, which removes the possibility of genomic integration of the TALEN-expressing protein. It can also dramatically increase the level of homology directed repair (HDR) and the success of introgression during gene editing.
Genome editing
TALENs can be used to edit genomes by inducing double-strand breaks (DSB), which cells respond to with repair mechanisms.
Non-homologous end joining (NHEJ) reconnects DNA from either side of a double-strand break where there is very little or no sequence overlap for annealing. This repair mechanism induces errors in the genome via insertion or deletion(indels), or chromosomal rearrangement; any such errors may render the gene products coded at that location non-functional.[8] Because this activity can vary depending on the species, cell type, target gene, and nuclease used, it should be monitored when designing new systems. A simple heteroduplex cleavage assay can be run which detects any difference between two alleles amplified by PCR. Cleavage products can be visualized on simple agarose gels or slab gel systems.
Alternatively, DNA can be introduced into a genome through NHEJ in the presence of exogenous double-stranded DNA fragments.[8]
Homology directed repair can also introduce foreign DNA at the DSB as the transfected double-stranded sequences are used as templates for the repair enzymes.[8]
TALENs have been used to generate stably modified human embryonic stem cell and induced pluripotent stem cell (IPSCs) clones,[9] to generate knockout C. elegans,[10] knockout rats,[11] knockout mice,[22] and knockout zebrafish.[12][23] Moreover, the method can be used to generate knockin organisms.Wu et al.obtain a Sp110 knockin cattle by Talen nickases to increased resistance of tuberculosis. [24] Also, the method has been used to generate Knockin rats by TALEN mRNA microinjection in one-cell embryos.[25]
TALENs have been utilized experimentally to correct genetic errors underlying disease.[26] TALENs have been used in vitro to correct the genetic defects that cause disorders such as sickle cell disease,[27] xeroderma pigmentosum,[28] and epidermolysis bullosa.[29] In theory, genome-wide specificity of engineered TALENs allows for correction of errors at individual genetic loci via homology-directed repair from a correct exogenous template.[26] In reality, however, in situ applications for TALENs are currently limited by the lack of an efficient delivery mechanism, unknown immunogenic factors, and uncertainty in the specificity of TALEN binding.[26]
Potential Problems
If a TAL effector nuclease is not specific enough for its target site or does not target a unique site within the genome of interest, off-target cleavage may occur. Such off-target cleavage may lead to the production of enough double-strand breaks to overwhelm the repair machinery and consequently yield chromosomal rearrangements and/or cell death.[16] Off-target cleavage events are possible with other forms of engineered nucleases, such as zinc finger nucleases, which have been more extensively researched for off-target activity.
See also
- Genome editing with engineered nucleases
- Zinc finger nuclease
- Meganuclease
- CRISPR
References
- ↑ 1.0 1.1 Boch, Jens (February 2011). "TALEs of genome targeting". Nature Biotechnology 29 (2): 135–6. doi:10.1038/nbt.1767. PMID 21301438.
- ↑ Boch, Jens et al. (December 2009). "Breaking the Code of DNA Binding Specificity of TAL-Type III Effectors". Science 326 (5959): 1509–12. Bibcode:2009Sci...326.1509B. doi:10.1126/science.1178811. PMID 19933107.
- ↑ Moscou, Matthew J.; Adam J. Bogdanove (December 2009). "A Simple Cipher Governs DNA Recognition by TAL Effectors". Science 326 (5959): 1501. Bibcode:2009Sci...326.1501M. doi:10.1126/science.1178817. PMID 19933106.
- ↑ Christian, Michelle et al. (October 2010). "Targeting DNA Double-Strand Breaks with TAL Effector Nucleases". Genetics 186 (2): 757–61. doi:10.1534/genetics.110.120717. PMC 2942870. PMID 20660643.
- ↑ Li, Ting et al. (August 2010). "TAL nucleases (TALNs): hybrid proteins composed of TAL effectors and FokI DNA-cleavage domain". Nucleic Acids Research 39: 1–14. doi:10.1093/nar/gkq704. PMC 3017587. PMID 20699274.
- ↑ Mahfouz, Magdy M. et al. (February 2010). "De novo-engineered transcription activator-like effector (TALE) hybrid nuclease with novel DNA binding specificity creates double-stran breaks". PNAS 108 (6): 2623–8. Bibcode:2011PNAS..108.2623M. doi:10.1073/pnas.1019533108. PMC 3038751. PMID 21262818.
- ↑ 7.0 7.1 7.2 Cermak, T.; Doyle, E. L.; Christian, M.; Wang, L.; Zhang, Y.; Schmidt, C.; Baller, J. A.; Somia, N. V.; Bogdanove, A. J.; Voytas, D. F. (2011). "Efficient design and assembly of custom TALEN and other TAL effector-based constructs for DNA targeting". Nucleic Acids Research 39 (12): e82. doi:10.1093/nar/gkr218. PMC 3130291. PMID 21493687.
- ↑ 8.0 8.1 8.2 8.3 8.4 Miller, Jeffrey et al. (February 2011). "A TALE nuclease architecture for efficient genome editing". Nature Biotechnology 29 (2): 143–8. doi:10.1038/nbt.1755. PMID 21179091.
- ↑ 9.0 9.1 9.2 Hockemeyer, D.; Wang, H.; Kiani, S.; Lai, C. S.; Gao, Q.; Cassady, J. P.; Cost, G. J.; Zhang, L.; Santiago, Y.; Miller, J. C.; Zeitler, B.; Cherone, J. M.; Meng, X.; Hinkley, S. J.; Rebar, E. J.; Gregory, P. D.; Urnov, F. D.; Jaenisch, R. (2011). "Genetic engineering of human pluripotent cells using TALE nucleases". Nature Biotechnology 29 (8): 731–734. doi:10.1038/nbt.1927. PMC 3152587. PMID 21738127.
- ↑ 10.0 10.1 Wood, A. J.; Lo, T. -W.; Zeitler, B.; Pickle, C. S.; Ralston, E. J.; Lee, A. H.; Amora, R.; Miller, J. C.; Leung, E.; Meng, X.; Zhang, L.; Rebar, E. J.; Gregory, P. D.; Urnov, F. D.; Meyer, B. J. (2011). "Targeted Genome Editing Across Species Using ZFNs and TALENs". Science 333 (6040): 307. doi:10.1126/science.1207773. PMC 3489282. PMID 21700836.
- ↑ 11.0 11.1 Tesson, L.; Usal, C.; Ménoret, S. V.; Leung, E.; Niles, B. J.; Remy, S. V.; Santiago, Y.; Vincent, A. I.; Meng, X.; Zhang, L.; Gregory, P. D.; Anegon, I.; Cost, G. J. (2011). "Knockout rats generated by embryo microinjection of TALENs". Nature Biotechnology 29 (8): 695–696. doi:10.1038/nbt.1940. PMID 21822240.
- ↑ 12.0 12.1 Huang, P.; Xiao, A.; Zhou, M.; Zhu, Z.; Lin, S.; Zhang, B. (2011). "Heritable gene targeting in zebrafish using customized TALENs". Nature Biotechnology 29 (8): 699. doi:10.1038/nbt.1939.
- ↑ Doyon, Y.; Vo, T. D.; Mendel, M. C.; Greenberg, S. G.; Wang, J.; Xia, D. F.; Miller, J. C.; Urnov, F. D.; Gregory, P. D.; Holmes, M. C. (2010). "Enhancing zinc-finger-nuclease activity with improved obligate heterodimeric architectures". Nature Methods 8 (1): 74–79. doi:10.1038/nmeth.1539. PMID 21131970.
- ↑ Szczepek, M.; Brondani, V.; Büchel, J.; Serrano, L.; Segal, D. J.; Cathomen, T. (2007). "Structure-based redesign of the dimerization interface reduces the toxicity of zinc-finger nucleases". Nature Biotechnology 25 (7): 786–793. doi:10.1038/nbt1317. PMID 17603476.
- ↑ Guo, J.; Gaj, T.; Barbas Iii, C. F. (2010). "Directed Evolution of an Enhanced and Highly Efficient FokI Cleavage Domain for Zinc Finger Nucleases". Journal of Molecular Biology 400 (1): 96–107. doi:10.1016/j.jmb.2010.04.060. PMC 2885538. PMID 20447404.
- ↑ 16.0 16.1 Mussolino, C.; Morbitzer, R.; Lutge, F.; Dannemann, N.; Lahaye, T.; Cathomen, T. (2011). "A novel TALE nuclease scaffold enables high genome editing activity in combination with low toxicity". Nucleic Acids Research. doi:10.1093/nar/gkr597.
- ↑ 17.0 17.1 Zhang, Feng et al. (February 2011). "Efficient construction of sequence-specific TAL effectors for modulating mammalian transcription". Nature Biotechnology 29 (2): 149–53. doi:10.1038/nbt.1775. PMC 3084533. PMID 21248753.
- ↑ Morbitzer, R.; Elsaesser, J.; Hausner, J.; Lahaye, T. (2011). "Assembly of custom TALE-type DNA binding domains by modular cloning". Nucleic Acids Research. doi:10.1093/nar/gkr151.
- ↑ Li, T.; Huang, S.; Zhao, X.; Wright, D. A.; Carpenter, S.; Spalding, M. H.; Weeks, D. P.; Yang, B. (2011). "Modularly assembled designer TAL effector nucleases for targeted gene knockout and gene replacement in eukaryotes". Nucleic Acids Research. doi:10.1093/nar/gkr188.
- ↑ Geiβler, R.; Scholze, H.; Hahn, S.; Streubel, J.; Bonas, U.; Behrens, S. E.; Boch, J. (2011). Shiu, Shin-Han, ed. "Transcriptional Activators of Human Genes with Programmable DNA-Specificity". PLoS ONE 6 (5): e19509. doi:10.1371/journal.pone.0019509.
- ↑ Weber, E.; Gruetzner, R.; Werner, S.; Engler, C.; Marillonnet, S. (2011). Bendahmane, Mohammed, ed. "Assembly of Designer TAL Effectors by Golden Gate Cloning". PLoS ONE 6 (5): e19722. doi:10.1371/journal.pone.0019722.
- ↑ Davies, B; Davies, G; Preece, C; Puliyadi, R; Szumska, D et al. (2013). "Site Specific Mutation of the Zic2 Locus by Microinjection of TALEN mRNA in Mouse CD1, C3H and C57BL/6J Oocytes". PLOS ONE 8: e60216. doi:10.1371/journal.pone.0060216.
- ↑ Sander et al. "Targeted gene disruption in somatic zebrafish cells using engineered TALENs". Nature Biotechnology 29: 697–98. doi:10.1038/nbt.1934.
- ↑ Wu et al. "TALE nickase-mediated SP110 knockin endows cattle with increased resistance to tuberculosis". Proceedings of the National Academy of Sciences of the United States of America 112: E1530–E1539. doi:10.1073/pnas.1421587112.
- ↑ Ponce; de León, V et al. (2014). ", Generation of TALEN-mediated GRdim knock-in rats by homologous recombination". PLOS ONE 9 (2): e88146. doi:10.1371/journal.pone.0088146.
- ↑ 26.0 26.1 26.2 Carlson, D. F. & Fahrenkrug, S. C. (2012). "Targeting DNA with fingers and TALENs" (PDF). Molecular Therapy- Nucleic Acids 1 (3): e3. doi:10.1038/mtna.2011.5. PMC 3381595. PMID 23344620. Retrieved November 11, 2014.
- ↑ Ramalingam, S. (2014). "TALEN-mediated generation and genetic correction of disease-specific human induced pluripotent stem cells." (URL). Current Gene Therapy 14: 461–472. doi:10.2174/1566523214666140918101725. Retrieved November 11, 2014.
- ↑ Dupuy, A. (2013). "Targeted gene therapy of xeroderma pigmentosum cells using meganuclease and TALEN™". PLOS ONE 8 (11): e78678. doi:10.1371/journal.pone.0078678. PMC 3827243. PMID 24236034.
- ↑ Osborn, M. J. (2013). "TALEN-based gene correction for epidermolysis bullosa" (URL). Molecular Therapy 21 (6): 1151–1159. doi:10.1038/mt.2013.56. Retrieved November 11, 2014.
External links
- E-TALEN.org A comprehensive tool for TALEN design
- TALengineering.org A comprehensive, publicly available resource for engineered TAL effector technology
- TALengineering newsgroup Newsgroup for discussion of engineered TAL effector technology
- www.taleffectors.com An open resource for TAL effector constructs
- PDB Molecule of the Month An entry in the Protein Database's monthly structural highlight