Somatic marker hypothesis
The somatic marker hypothesis (SMH) proposes a mechanism by which emotional processes can guide (or bias) behavior, particularly decision-making.[1][2] This hypothesis has been formulated by Antonio Damasio.
Hypothesis
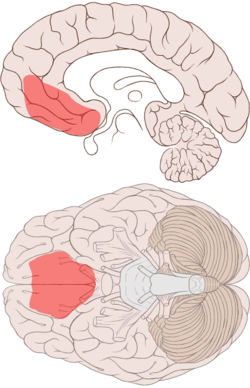
When individuals make decisions, they must assess the incentive value of the choices available to them, using cognitive and emotional processes. When the individuals face complex and conflicting choices, they may be unable to decide using only cognitive processes, which may become overloaded.
In these cases (and others), somatic markers can help decide. Somatic markers are associations between reinforcing stimuli that induce an associated physiological affective state. Within the brain, somatic markers are thought to be processed in the ventromedial prefrontal cortex (VMPFC; a subsection of the orbitomedial PFC, OMPFC).[3] These somatic marker associations can recur during decision-making and bias one's cognitive processing. When one has to make complex and uncertain decisions, the somatic markers created by the relevant stimuli are summed to produce a net somatic state. This overall state directs (or biases) one's decision of how to act.[4] This influence on our decision-making process may occur covertly (unconsciously), via the brainstem and ventral striatum, or overtly (consciously), engaging higher cortical cognitive processing. Damasio proposes that somatic markers direct attention towards more advantageous options, simplifying the decision process.
The amygdala and VMPFC are essential components of this hypothesized mechanism and therefore damage to either structure will disrupt their proposed action in mediating the development and action of somatic markers. A major source of supporting evidence for this theory is provided by experiments using the Iowa gambling task.[5]
Research background
In economic theory, there is a tendency to model human decision-making as being devoid of emotions, involving only logical reasoning based on cost-benefit calculations.[6] Such theories assume that individuals have the time, knowledge and information processing power to make optimal decisions. In contrast to this idealization, the somatic marker hypothesis proposes that emotions play a critical role in the ability to make fast, rational decisions in complex and uncertain situations.[1]
Patients with frontal lobe damage (e.g., Phineas Gage) provided the first evidence that the frontal lobes were associated with decision-making and social conduct. Frontal lobe damage, particularly to the VMPFC, results in impaired abilities to organize and plan behavior, learn from previous mistakes, and behave in a socially appropriate manner.[7] Patients with damage to the VMPFC develop severe impairments in both personal and social decision-making that can result in choosing unsuitable friends, partners, and activities. In fact, these patients appear to lack concern for other individuals, which resembles a mild form of sociopathy.[8] Surprisingly, patients with VMPFC damage have normal intellect in terms of working memory, attention, and language comprehension and expression.
VMPFC patients also have difficulty expressing and experiencing appropriate emotions. This led Antonio Damasio to hypothesize that decision-making deficits following VMPFC damage result from the inability to use emotions to help guide future behavior based on past experiences. This damage prevents rapid emotional signaling to bias behaviors toward appropriate responses to simplify the process. Consequently, VMPFC damage forces those afflicted to rely on slow and laborious cost-benefit analyses for every given choice situation,[9] which degrades accuracy and response-time.
Working mechanism
Emotions, as defined by Damasio, are changes in both body and brain states in response to different stimuli.[1] Physiological changes (e.g., muscle tone, heart rate, endocrine release, posture, facial expression, etc.) occur in the body and are relayed to the brain where they are transformed into an emotion that tells the individual something about the stimulus that they have encountered. Over time, emotions and their corresponding bodily change(s) become associated with particular situations and their past outcomes.
When making decisions, these physiological signals (or ‘somatic markers’) and their evoked emotion are consciously or unconsciously associated with their past outcomes and bias decision-making towards certain behaviors while avoiding others.[1] For instance, when a somatic marker associated with a positive outcome is perceived, the person may feel happy and thereby motivated to pursue that behavior. When a somatic marker associated with the negative outcome is perceived, the person may feel sad, which feeling acts as an internal alarm to warn the individual to avoid a course of action. These situation-specific somatic states based on, and reinforced by, past experiences help to guide behavior in favor of more advantageous choices and therefore are adaptive.
According to the SMH, two distinct pathways reactivate somatic marker responses. In the first pathway, emotion can be evoked by the changes in the body that are projected to the brain—called the “body loop”. For instance, encountering a feared object like a snake may initiate the fight-or-flight response and cause fear. In the second pathway, cognitive representations of the emotions can be activated in the brain without being directly elicited by a physiological response—called the “as-if body loop”. For instance, imagining an encounter with a snake would initiate a similar flight-or-fight response “as-if” you were in that particular situation (albeit perhaps a much weaker one). In other words, the brain can anticipate expected bodily changes, which allows the individual to respond faster to external stimuli without waiting for an event to actually occur.[7]
Evolutionary evidence
To support his claims about the evolution of the human race, Charles Darwin wrote The Expression of Emotions in Man and Animals in which he noted the similarities between emotional facial expressions between humans and other animals. In this book, Darwin argues that human emotion-evoked expressions, like furrowing of the eyebrows and tears, are similar to animal facial expressions and is evidence of our evolutionary origins. Today, the existence of emotion in animals is controversial, but current research suggests that intelligent, social animals (e.g., primates, dolphins, parrots)[10] do experience primary emotions like fear, joy, anger, and disgust.[11] In particular, our closest living relatives, the great apes, are prime candidates for having the most developed capabilities for emotions, empathy, and theories of mind.
Within an evolutionary framework, emotions in humans and animals can be defined as coordinated cognitive and physiological responses that enable an individual to respond effectively to environmental opportunities and communicate with others.[10] For example, anger is not simply a specific facial expression or neural activation, rather it is a set of coordinated responses that help an individual express dissatisfaction and perhaps restore relations. Emotions are adaptive because they are efficient responses that help organisms reproduce, protect offspring, maintain cooperative alliances, and avoid physical dangers.[12] Emotions and their influence on motivation and certain physiological responses, prepare individuals for actions that are in their best interest. In humans, for example, anger is associated with enhanced blood flow to the hands for fighting an enemy, whereas fear causes less blood flow to the periphery to help escape an attack without too much blood loss.[13]
Yet humans have a much larger array of finely tuned emotions compared to animals, even the great apes. It has been hypothesized that concurrent selection for greater social cohesion and organizational flexibility in early hominids may have been the impetus for increasing the human emotional repertoire.[14] The elaboration of human emotions may have aided social bonds if genetic relatedness was low within a social group. Consequently, the increase in the spectrum of human emotions could also be used to tag dimensions within their environment with emotional value that could be used in the future to guide difficult decisions that had not yet been encountered by animals— such as the abstract and complex issues surrounding morals and social rules.
According to the Social brain hypothesis the demand for the ability to solve complex social problems initiated the evolutionary trend for increases in brain size observed among humans and other primates.[15][16] This increase is mostly due to an increase in prefrontal cortex volume. The evolution of the human prefrontal cortex allows humans to think and process information in more abstract ways. By tagging particular stimuli with an emotion associated with a bodily change, it helps to speed up the process of decision-making when it is encountered by eliminating unsuitable choices and ensuring only advantageous options are considered and fully processed. This is most important in the social domain since social environments are complex, unpredictable and more responsive to one’s own behavior than the physical environment. Since social interactions are complex, evaluating the appropriate behavioral response in these situations requires greater brain processing capacity involving multiple brain structures.[17] Given that there is a large amount of information that needs to be processed in a short amount of time, it is advantageous to have a “fast-track decision-making” mechanism (i.e., somatic markers), that can respond rapidly in the most advantageous fashion. In essence, the SMH provides neurobiological evidence of what has often been referred to as “hunches” or “gut-feelings”.[9]
The SMH is supported by human lesion studies suggesting that predictions of future outcomes or consequences that are near in time require processing in the more posterior portion of the ventromedial (VM) cortices and representations of outcomes or consequences not in the immediate future are processed in the anterior portion of the VM cortices.[9] Patients with damage to only the anterior VM cortices, are more likely to engage in behaviors that negatively impact personal relationships in the distant future. However, they will never engage in actions that would lead to immediate harm to themselves or others.[1] This brain organization pattern appears to be rooted in evolution. The functional evolution of the prefrontal cortex involves increases in the ability to represent events that may occur in the future.[9] The ability for humans to think about future consequences of their actions coincides with the development of the more anterior portions of the VM cortices.[18][19] This is also supported by human neuroimaging studies demonstrating that tasks involving moral or ethical decisions evoke increased activity in the most anterior sections of the VM cortex.[20] Finally, comparative studies in neuroanatomy demonstrate that advancements in the size and connectivity seen in human brains primarily relates to the evolutionarily younger anterior portions of the frontal lobes as opposed to the more ‘ancient’ posterior areas.[18][19] Consequently, Damasio suggests that the ability of humans to cope with such degrees of abstract thinking quickly and efficiently coincides with both the development of the anterior regions of the VM cortex and the use of somatic markers to guide human behavior.[9]
Experiments
In a quest to produce a simple neuropsychological tool that would adequately assess the obvious deficits in emotional processing, decision-making, and social skills of OMPFC lesioned individuals[2] Bechara et al. 1994[5] created the Iowa gambling task. Their aim was to produce "[a] task which simulates in real time, personal real-life decision-making relative to the way it factors uncertainty of premises and outcomes, as well as reward and punishment" (p. 8). The task measures a form of learning that has been defined as emotion-based learning. Studies using the gambling task have found deficits in various neurological (e.g. amygdala and OMPFC) and psychiatric populations (e.g. schizophrenia, mania, drug abusers), providing support for the SMH.
Iowa Gambling Task
The human brain has evolved over time to best benefit one's genes in any decision-making circumstance.[21] It is important to understand that “natural selection shapes not only the physical characteristics of organisms, but also their behavioral and cognitive traits”.[22]
The Iowa Gambling Task is the most common experimental paradigm used to test decision-making processes under various contexts and is frequently used in experiments exploring the Somatic Marker Hypothesis. The Iowa Gambling Task is a computerized task in which participants are presented with four decks of cards from which they repeatedly choose. Each deck contains various amounts of rewards of either $50 or $100, and occasional losses that are greater in the decks with higher rewards. The penalty cards are periodically located in the deck so the participant will not know when they will arise and the participants are told to pick cards that will maximize their winnings. The most profitable strategy is to only choose cards from the small reward/penalty decks because although the reward is smaller, the penalty is proportionally much smaller than in the higher reward/penalty deck. Over the course of a session, this deck will yield a significantly higher net profit compared to the alternative deck. Most healthy control participants adopt the beneficial low reward/penalty deck strategy during the duration of the task. Participants with brain damage however, are unable to determine the better deck to choose from, and continue to choose from the high reward/penalty deck despite this action causing a clear lower net profit than the other deck.[23] This experiment has been used to analyze the impairments suffered by people with damage to the ventromedial prefrontal cortex, which has been known to affect neural signaling of prospective rewards or punishments.[1] Since the Iowa Gambling Task measures participants’ quickness in “developing anticipatory emotional responses to guide advantageous choices,” it is helpful in testing the Somatic Marker Hypothesis since it studies how anticipatory signals affect decision-making.[24] The study of human behavior in respect to evolutionary psychology demonstrates “that much, if not all, of our behavior can be explained by appeal to internal psychological mechanisms”.[25] The Iowa Gambling Task and the Somatic Marker Hypothesis relate to this theory by revealing that emotions may have evolved during the course of human evolution to help people make better decisions. Therefore, the brain has been trained to make immediate decisions that will benefit the decision-maker. Evolutionary theory and the Somatic Marker Hypothesis suggest that human emotions have evolved to send signals to the brain which will help someone make quick decisions that will benefit them. The Iowa Gambling Task upholds this evolutionary theory by demonstrating that lesions to the ventromedial prefrontal cortex act as a blockade to the emotional decision-making signals that have evolved in helping humans make quick and beneficial decisions. As suggested by Comsides and Tooby, “Mechanisms involved in hierarchically ranking goals or calibrating other kinds of motivational and reward systems should be emotion-dependent”.[26]
Although the Iowa Gambling Task is a well-known experimental measure used to explore how emotions may have evolved in humans to guide decision-making, there are “real-life” factors that can be added to the Iowa Gambling Task to test differences in participants’ results. One such study found that people who were given more time to complete the Iowa Gambling Task had better results compared to people that had less time [27] In a more recent study aimed to determine the effects of perceived time constraints, the experimental group was told that they would likely not be able to finish the task in the time allotted, whereas the control group was informed they had a sufficient amount of time to finish. As expected, the participants who were informed they had an insufficient amount of time to complete the task performed more poorly than the participants who were told they had ample time to finish it. The results of this test can be correlated to the real world in which professionals are told they have an adequate amount of time to complete a project are less likely to make mistakes.[28] Thus, recent studies suggest that the Iowa Gambling Task does not appropriately mimic “real-life” decision-making tasks because other factors, such as time, play into one’s success in the task instead of merely the health of the ventromedial prefontal cortex. However, human neuroimaging studies support the validity of the Iowa Gambling Task and show a clear link to “real-life” decision-making situations. Li and colleagues (2010) used functional magnetic resonance imaging (fMRI) to analyze the brain during the Iowa Gambling Task. This imaging reveals brain activity during the Iowa Gambling Task and indicates which parts of the brain are being used during decision-making processes. The results suggest that the brain regions that were activated during the Iowa Gambling Task were consistent with the ones hypothesized to trigger decision-making by somatic markers (i.e. brain regions involved in emotional processing). This adds validity to the Iowa Gambling task in conjunction with the Somatic Marker Hypothesis and suggests emotional processing is causing these regions to be activated as well as regions associated with decision-making.[29]
Application To Risky Sexual Behavior
Emotional decision-making can greatly affect aspects of people’s daily lives, such as their sex drive. Sensation is connected to the brain and likely stimulates precarious sexual behavior by making the riskier sexual behaviors more exhilarating and pleasurable.[30] The risky sexual behavior evaluated in a study by Wardle and colleagues was continued sexual activity in individuals who are already infected with HIV and are substance dependent. The Somatic Marker Hypothesis proposes that the Iowa Gambling Task may distinguish HIV+ and substance dependent people who have emotional influenced risks, from those who have risks caused by other, unrelated factors. The Somatic Marker Hypothesis was tested in a circumstance when extraneous factors become present, such as a lethal sexually transmitted disease and substance abuse. Wardle and colleagues performed a study utilizing the Iowa Gambling Task in which they drew together 190 HIV+ participants in the Chicago area who all had a history of drug dependency or abuse. Among the factors that were held constant in all participants were: education, race, and brain related items such as no history of neurological disorders or head injury. It was hypothesized that the Iowa Gambling Task would reveal that HIV+ substance dependent people, who are at increased risk for impairment and emotional suffering, are “motivated by negative emotion in their sexual risks”.[24] The results of the study supported the hypothesis that with the better performers on the Iowa Gambling Task there was a clear connection between distress and risk relatedness. The greater the distress, the greater risk these people would take in regards to sexual acts. However, the poor performers on the Iowa Gambling Task did not show a significant link between distress and sexual risk. These findings suggest that people with intact decision-making abilities can attenuate their risk-seeking behavior by decreasing their emotional distress. This conclusion is inconsistent with the Somatic Marker Hypothesis that posits that people with “dysfunctional decision-making circuitry as reflected by poor Iowa Gambling Task performance and emotional distress has little influence on their decision-making capacity”.[24] Instead of concluding that the Iowa Gambling Task is not fully accurate in demonstrating how we have psychologically evolved, it can be determined that sexual risk is a complex neurocognitive process, and emotional variables do not have as much weight in this aspect as in others actions by humans. Additionally, the entirety of human evolutionary psychology evolved without the implications of modern day drugs such as cocaine and other similar substances that have only been presently used and distributed for the past several hundred years.[31] Therefore, it is highly likely that the new initiation of drugs and also diseases (which have always been present through evolution) act as a similar blockade as a lesion to the ventromedial prefrontal cortex, which affects neural signaling. Though many “Theories about innate human predispositions are extremely difficult to verify” the Somatic Marker Hypothesis has clear evidence leading to its validity as tested in the Iowa Gambling Task.[32]
Drug Addiction
Despite being aware of the medical, social and legal problems associated with consuming illegal substances, substance dependent individuals (SDI) incessantly take part in activities that ultimately lead to addiction and dependency. This myopia for the future is characteristic of drug abusers and can be applied to somatic markers.[33] The Somatic Marker Hypothesis attributes SDI difficulty in making advantageous decisions to a defect in an emotional mechanism, which indicates the future consequences of an action and helps select the best response. This emotional mechanism is a special feeling that emerges in bioregulatory processes, and can be produced in the body or in brain areas.[34] When a negative somatic marker (like fear) is juxtaposed to a particular future outcome, it functions like a warning, signaling us to refrain from an action. Positive somatic markers act as an added incentive to behave in a particular way. Through this process, somatic markers help control human tendency to discount the future since long term costs associated with a negative somatic marker have the potential to deter an individual away from making a decision.[1]
According to the SMH, there should be a connection between abnormalities in expressing emotions and experiencing feelings, and severe impairments in decision-making. Much of the evidence for this comes from the Iowa Gambling Task (IGT), which provides testing of human decision processes in regard to immediate rewards and delayed punishments, risks, and uncertainty of outcomes.[1] The results from the IGT studies support the notion that impaired decision-making in substance dependent individuals is associated with varying reactions to rewarding and punishing events.[33] For example, it was apparent which packs of cards had the greatest risk for loss, but individuals continued to choose from the high-risk, high-reward packs. Thus the prospect of immediate high reward clearly outweighed the negative long term consequences for SDI. Damasio (1994) suggests that somatic markers provide a covert, non-conscious estimate of which cards are good and bad based on the rewards and punishments received. Prior to any conscious cognitive process of selection, initial sorting occurs and then the individual is guided in a theory about the gambling game more efficiently. Somatic markers, put more simply, appear to be a fast mechanism for reasoning that allows individuals to make satisfactory decisions without the necessary time to go through a lengthy analysis. They are advantageous, and thus adaptive.[1]
One of the more frequently used models, the International Affective Picture System (IAPS), consists of a large set of images categorized according to their normative values in three dimensions: valence, arousal, and control.[34] Using this model, Gerra et al. (2003) analyzed the neuroendocrine responses of both substance dependent individuals and healthy individuals in order to induce pleasant and unpleasant emotions. The results showed that in response to unpleasant images, SDI showed decreased activity in several neuroendocrine markers, including norepinephrine, cortisol, and adrenocorticotropic hormone levels. SDI showed a more level response pattern to both pleasant and unpleasant images, suggesting that they may have a diminished emotional response to natural reinforcers other than drugs.[35]
Consistent with this evidence, a neuroimaging study conducted on drug craving by Garavan et al. (2000) demonstrates that drug related stimuli have the ability to activate brain regions involved in emotional evaluation and reward processing. This study exposed two groups, one including experienced cocaine users (N=17) and the other including non-users (N=14), to three separate films: individuals smoking crack cocaine, outdoor nature scenes, and explicit sexual content. Meanwhile, all patients underwent functional magnetic resonance imaging, which monitored thirteen regions of the brain. Three regions of the brain, the anterior cingulated, right inferior parietal lobule, and the caudate/lateral dorsal nucleus, displayed significantly greater activation during the cocaine film than during the sex film in the cocaine users, suggesting that cocaine cues activate similar parts of the brain as natural stimuli in cocaine users. Additionally, cocaine users exhibited a lower response than the non-users to the sex film, suggesting that drug-users demonstrate a lower emotional response to natural reinforcers other than drugs.[36] Although the somatic states linked with natural reinforcers may not be strong enough to influence decisions in substance dependent individuals, these studies demonstrate how strong somatic states associated with drug abuse have the potential to dominate decisions regarding drug use. Modern day drugs like cocaine were not popular throughout human evolution, and so it seems obvious that natural responses are affected by habitual cocaine use.[34]
Criticism
Some researchers believe that the use of somatic markers (i.e., afferent feedback) would be a very inefficient method of influencing behavior. Damasio's notion of the as-if experience dependent feedback route,[1][4] whereby bodily responses are re-represented utilizing the somatosensory cortex (postcentral gyrus), also proposes an inefficient method of affecting explicit behavior.[37] Rolls (1999) stated that; "it would be very inefficient and noisy to place in the execution route a peripheral response, and transducers to attempt to measure that peripheral response, itself a notoriously difficult procedure" (p. 73).[37] Reinforcement association located in the orbitofrontal cortex and amygdala, where the incentive value of stimuli is decoded, is sufficient to elicit emotion-based learning and to affect behavior via, for example, the orbitofrontal-striatal pathway.[37][38] This process can occur via implicit or explicit processes.[37]
The Somatic Marker Hypothesis represents an intriguing model of how feedback from the body may contribute to both advantageous and disadvantageous decision-making in situations of complexity and uncertainty. Much of this support comes from data taken from the Iowa Gambling Task.[39] While the Iowa Gambling Task has proven to be an ecologically valid measure of decision-making impairment, there exist three assumptions that need to hold true. First, the claim that it assesses implicit learning as the reward/punishment design is inconsistent with data showing accurate knowledge of the task possibilities [40] and that mechanisms such as working-memory appear to have a strong influence. Second, the claim that this knowledge occurs through preventive marker signals is not supported by competing explanations of the psychophysiology generated profile.[41] Lastly, the claim that the impairment is due to a ‘myopia for the future’ is undermined by more plausible psychological mechanisms explaining deficits on the tasks such as reversal learning, risk-taking, and working-memory deficits. There may also be more variability in control performance than previously thought, thus complicating the interpretation of the findings. Furthermore, although the Somatic Marker Hypothesis has accurately identified many of the brain regions involved in decision-making, emotion, and body-state representation, it has failed to clearly demonstrate how these processes interact at a psychological and evolutionary level. There are many experiments that could be implemented to further test the Somatic Marker Hypothesis. One way would be to develop variants of the Iowa Gambling Task that control some of the methodological issues and interpretation ambiguities generated. It may be a good idea to include removing the reversal learning confound, which would make the task more difficult to consciously comprehend. Additionally, causal tests of the Somatic Marker Hypothesis could be practiced more insistently in a greater range of populations with altered peripheral feedback, like on patients with facial paralysis. In conclusion, the Somatic Marker Hypothesis needs to be tested in more experiments. Until a wider range of empirical approaches are employed in order to test the Somatic Marker Hypothesis, it appears that the framework is simply an intriguing idea that is in need of some better supporting evidence. Despite these issues, the Somatic Marker Hypothesis and the Iowa Gambling Task reestablish the notion that emotion has the potential to be a benefit as well as a problem during the decision-making process in humans.[42]
References
- ↑ 1.0 1.1 1.2 1.3 1.4 1.5 1.6 1.7 1.8 1.9 Damasio, Antonio R. (2008) [1994]. Descartes' Error: Emotion, Reason and the Human Brain. Random House. ISBN 978-1-4070-7206-7. Descartes' Error
- ↑ 2.0 2.1 Damasio, A.R.; Tranel, D.; Damasio, H.C. (1991). "Ch. 11: Somatic markers and the guidance of behaviour: theory and preliminary testing". In Levin, Harvey S.; Eisenberg, Howard M.; Benton, Arthur Lester. Frontal Lobe Function and Dysfunction. Oxford University Press. pp. 217–229. ISBN 978-0-19-506284-7.
- ↑ Damasio, A.R.; Everitt, B. J.; Bishop, D. (29 October 1996). "The Somatic marker hypothesis and the possible functions of the prefrontal cortex". Phil. Trans. R. Soc. Lond. B 351 (1346): 1413–20. doi:10.1098/rstb.1996.0125. PMID 8941953.
- ↑ 4.0 4.1 Damasio, Antonio R. (2000). The Feeling of what Happens: Body, Emotion and the Making of Consciousness. Vintage. ISBN 978-0-09-928876-3.
- ↑ 5.0 5.1 Bechara A, Damasio AR, Damasio H, Anderson SW; Damasio; Damasio; Anderson (1994). "Insensitivity to future consequences following damage to human prefrontal cortex". Cognition 50 (1–3): 7–15. doi:10.1016/0010-0277(94)90018-3. PMID 8039375.
- ↑ Loewenstein GF, Weber EU, Hsee CK, Welch N; Weber; Hsee; Welch (March 2001). "Risk as feelings". Psychol Bull 127 (2): 267–86. doi:10.1037/0033-2909.127.2.267. PMID 11316014.
- ↑ 7.0 7.1 Damasio, A. (1991). Somatic Markers and the Guidance of Behavior. New York: Oxford University Press. pp. 217–299.
- ↑ Tranel D (1994). ""Acquired sociopathy": the development of sociopathic behavior following focal brain damage". Prog Exp Pers Psychopathol Res: 285–311. PMID 8044207.
- ↑ 9.0 9.1 9.2 9.3 9.4 Bechara, A.; Damasio, A.R. (2005). "The somatic marker hypothesis: A neural theory of economic decision". Games and Economic Behavior 52 (2): 336–372. doi:10.1016/j.geb.2004.06.010.
- ↑ 10.0 10.1 Alexander, R.D. (1989). Mellars, Paul; Stringer, Christopher, eds. The Human Revolution: Behavioural and Biological Perspectives on the Origins of Modern Humans. Princeton NJ: Princeton University Press.
- ↑ Bekoff, M. (2000). "Animal emotions: Exploring Passionate Natures". BioScience 50 (10): 861–870. doi:10.1641/0006-3568(2000)050[0861:AEEPN]2.0.CO;2.
- ↑ Tooby, C.; Cosmides, L. (1990). "The past explains the present: Emotional adaptations and the structure of ancestral environments". Ethology and Sociobiology 11 (4–5): 375–424. doi:10.1016/0162-3095(90)90017-Z.
- ↑ Levenson, R.W. (1992). "Autonomic nervous system differences among emotions". Psychological Sciences 3: 23–27. doi:10.1111/j.1467-9280.1992.tb00251.x.
- ↑ Turner, J.H. (1996). "The evolution of emotions in humans: A Darwinian-Durkheimian analysis". Journal for the Theory of Social Behavior 26: 1–33. doi:10.1111/j.1468-5914.1996.tb00283.x.
- ↑ Humphrey, N.K. (1976). "Ch. 9: The social function of intellect". In Bateson, Paul Patrick Gordon; Hinde, R.A. Growing Points Ethology. CUP Archive. pp. 303–317. ISBN 978-0-521-29086-9.
- ↑ Dunbar, R. (1998). "The social brain hypothesis". Evolutionary Anthropology 6 (5): 178–190. doi:10.1002/(SICI)1520-6505(1998)6:5<178::AID-EVAN5>3.0.CO;2-8.
- ↑ Adolphs R (April 2001). "The neurobiology of social cognition". Curr. Opin. Neurobiol. 11 (2): 231–9. doi:10.1016/S0959-4388(00)00202-6. PMID 11301245.
- ↑ 18.0 18.1 Semendeferi K, Armstrong E, Schleicher A, Zilles K, Van Hoesen GW; Armstrong; Schleicher; Zilles; Van Hoesen (March 2001). <224::AID-AJPA1022>3.0.CO;2-I "Prefrontal cortex in humans and apes: a comparative study of area 10". Am. J. Phys. Anthropol. 114 (3): 224–41. doi:10.1002/1096-8644(200103)114:3<224::AID-AJPA1022>3.0.CO;2-I. PMID 11241188.
- ↑ 19.0 19.1 Semendeferi, K. et al. (2002). "Humans and great apes share a large frontal cortex". Nature Neuroscience 5 (3): 272–276. doi:10.1038/nn814. PMID 11850633.
- ↑ Greene, J.D. et al. (2001). "An fMRI investigation of emotional engagement in moral judgement". Science 293 (5537): 2105–2108. doi:10.1126/science.1062872. PMID 11557895.
- ↑ Dawkins, Richard (2006). The Selfish Gene (30th Anniversary edition). New York City: Oxford University Press. ISBN 0-19-929115-2.
- ↑ Durrant, Russil, and Bruce Ellis. "Evolutionary Psychology." psychology 1 (2002): 1-34. Print. Go, Pauline. "Where Does Cocaine Come From." Ezine 1 (2011): 1-2. Print
- ↑ Dezfouli AK, Keramati MM, Ekhtiari H, Safaei H, Lucas C (2008). "Understanding Addictive Behavior on the Iowa Gambling Task Using Reinforcement Learning Framework". Proceedings of the 30th Annual Conference of the Cognitive Science Society. pp. 1094–9.
- ↑ 24.0 24.1 24.2 Wardle MC, Gonzalez R, Bechara A, Martin-Thormeyer EM; Gonzalez; Bechara; Martin-Thormeyer (December 2010). "Iowa Gambling Task performance and emotional distress interact to predict risky sexual behavior in individuals with dual substance and HIV diagnoses". J Clin Exp Neuropsychol 32 (10): 1110–21. doi:10.1080/13803391003757833. PMC 3639122. PMID 20480423.
- ↑ Downes, Stephen (8 February 2008). "Evolutionary Psychology". Stanford Encyclopedia of Philosophy. Metaphysics Research Lab, CSLI, Stanford University.
- ↑ Cosmides, Leda; Tooby, John (2000). "Ch. 7: Evolutionary Psychology and the Emotions". In Lewis, Michael; Haviland-Jones, Jeannette M. Handbook of Emotions (2nd ed.). Guilford. pp. 91–115. ISBN 1572305290.
- ↑ Cella M, Dymond S, Cooper A, Turnbull O; Dymond; Cooper; Turnbull (July 2007). "Effects of decision-phase time constraints on emotion-based learning in the Iowa Gambling Task". Brain Cogn 64 (2): 164–9. doi:10.1016/j.bandc.2007.02.003. PMID 17368682.
- ↑ DeDonno MA, Demaree HA (December 2008). "Perceived Time Pressure and the Iowa Gambling Task". Judgment and Decision Making 31 (8): 636–640.
- ↑ Li X, Lu ZL, D'Argembeau A, Ng M, Bechara A; Lu; d'Argembeau; Ng; Bechara (March 2010). "The Iowa Gambling Task in fMRI images". Hum Brain Mapp 31 (3): 410–23. doi:10.1002/hbm.20875. PMC 2826566. PMID 19777556.
- ↑ Zuckerman M, Kuhlman DM; Kuhlman (December 2000). "Personality and risk-taking: common biosocial factors". J Pers 68 (6): 999–1029. doi:10.1111/1467-6494.00124. PMID 11130742.
- ↑ Go, Pauline. "Where Does Cocaine Come From." Ezine 1 (2011): 1-2. Print.
- ↑ Harris CR, Pashler HE; Pashler (1995). "Evolution and Human Emotions" (PDF). Psychological Inquiry 6 (1): 44–46. doi:10.1207/s15327965pli0601_6. JSTOR 1449573.
- ↑ 33.0 33.1 Bechara A (2003). "Risky business: emotion, decision-making, and addiction". J Gambl Stud 19 (1): 23–51. doi:10.1023/A:1021223113233. PMID 12635539.
- ↑ 34.0 34.1 34.2 Verdejo-García A, Bechara A, Recknor EC, Pérez-García M; Bechara; Recknor; Pérez-García (May 2006). "Executive dysfunction in substance dependent individuals during drug use and abstinence: an examination of the behavioral, cognitive and emotional correlates of addiction". J Int Neuropsychol Soc 12 (3): 405–15. doi:10.1017/S1355617706060486. PMID 16903133.
- ↑ Gerra G, Baldaro B, Zaimovic A et al. (July 2003). "Neuroendocrine responses to experimentally-induced emotions among abstinent opioid-dependent subjects". Drug Alcohol Depend 71 (1): 25–35. doi:10.1016/S0376-8716(03)00065-6. PMID 12821203.
- ↑ Garavan H, Pankiewicz J, Bloom A et al. (November 2000). "Cue-induced cocaine craving: neuroanatomical specificity for drug users and drug stimuli". Am J Psychiatry 157 (11): 1789–98. doi:10.1176/appi.ajp.157.11.1789. PMID 11058476.
- ↑ 37.0 37.1 37.2 37.3 (). Rolls, E.T. (1999). The brain and emotion. Oxford: Oxford University Press. ISBN 978-0-19-852464-9.
- ↑ Rolls ET (October 1997). "Consciousness in Neural Networks?". Neural Netw 10 (7): 1227–40. doi:10.1016/S0893-6080(97)00049-X. PMID 12662513.
- ↑ Bechara A, Tranel D, Damasio H, Damasio AR; Tranel; Damasio; Damasio (1996). "Failure to respond autonomically to anticipated future outcomes following damage to prefrontal cortex". Cereb. Cortex 6 (2): 215–25. doi:10.1093/cercor/6.2.215. PMID 8670652.
- ↑ Maia TV, McClelland JL; McClelland (November 2004). "A reexamination of the evidence for the somatic marker hypothesis: what participants really know in the Iowa gambling task". Proc. Natl. Acad. Sci. U.S.A. 101 (45): 16075–80. doi:10.1073/pnas.0406666101. PMC 528759. PMID 15501919.
- ↑ Tomb I, Hauser M, Deldin P, Caramazza A; Hauser; Deldin; Caramazza (November 2002). "Do somatic markers mediate decisions on the gambling task?". Nat. Neurosci. 5 (11): 1103–4; author reply 1104. doi:10.1038/nn1102-1103. PMID 12403997.
- ↑ Dunn BD, Dalgleish T, Lawrence AD; Dalgleish; Lawrence (2006). "The somatic marker hypothesis: a critical evaluation". Neurosci Biobehav Rev 30 (2): 239–71. doi:10.1016/j.neubiorev.2005.07.001. PMID 16197997.
External links
- Bechara A, Damasio H, Damasio AR; Damasio; Damasio (March 2000). "Emotion, decision making and the orbitofrontal cortex". Cereb. Cortex 10 (3): 295–307. doi:10.1093/cercor/10.3.295. PMID 10731224.