Silent synapse
In neuroscience, a silent synapse is an excitatory glutamatergic synapse whose postsynaptic membrane contains NMDA-type glutamate receptors but no AMPA-type glutamate receptors.[1] These synapses are named "silent" because normal AMPA receptor-mediated signaling is not present, rendering the synapse inactive under typical conditions. Silent synapses are typically considered to be immature glutamatergic synapses. As the brain matures, the relative number of silent synapses decreases. However, recent research on hippocampal silent synapses shows that while they may indeed be a developmental landmark in the formation of a synapse, that synapses can be "silenced" by activity, even once they have acquired AMPA receptors. Thus, silence may be a state that synapses can visit many times during their lifetimes.
Synaptic transmission
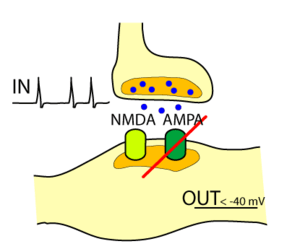
Normal transmission across a glutamatergic synapse relies on the neurotransmitter glutamate, the glutamate-specific AMPA receptor (AMPAR), and calcium ions. Calcium ion entry into the presynaptic terminal causes the presynaptic release of glutamate, which diffuses across the synaptic cleft, binding to glutamate receptors on the postsynaptic membrane. There are four subtypes of glutamate receptors: AMPA receptors (AMPARs) (formerly known as quisqualate receptors), NMDA receptors (NMDARs), kainate receptors, and metabotropic glutamate receptors (mGluRs). Most research has been focused on the AMPARs and the NMDARs. When glutamate binds to AMPARs located on the postsynaptic membrane, they permit a mixed flow of Na+ and K+ to cross the cells membrane, causing a depolarization of the postsynaptic membrane. This localized depolarization is called an excitatory postsynaptic potential (EPSP).
Silent synapses release glutamate as do prototypical glutamatergic synapses, but their postsynaptic membranes contain only NMDA—and possibly mGlu—receptors able to bind glutamate. Though AMPA receptors are not expressed in the postsynaptic membranes of silent synapses, they are stored in vesicles inside the postsynaptic cells, where they cannot detect extracellular glutamate, but can be quickly inserted into the postsynaptic cell membrane in response to a tetanizing stimulus. The NMDAR is functionally similar to AMPAR except for two major differences: NMDARs carry ion currents composed of Na+, K+, but also (unlike most AMPAR) Ca2+; NMDARs also have a site inside their ion channel that binds magnesium ions (Mg2+). This magnesium binding site is located in the pore of the channel, at a place within the electrical field generated by the membrane potential. Normally, current will not flow through the NMDAR channel, even when it has bound glutamate. This is because the ion channel associated with this receptor is plugged by magnesium, acting like a cork in a bottle. However, since the Mg2+ is charged and is bound within the membrane's electric field, depolarization of the membrane potential above threshold can dislodge the magnesium, allowing current flow through the NMDAR channel. This gives the NMDAR the property of being voltage-dependent, in that it requires strong postsynaptic depolarization to allow ion flux.
Characteristics
The stimulation of a silent synapse does not elicit EPSPs when the postsynaptic cell is clamped at -60 mV. Stimulation of a silent synapse will elicit EPSPs when the postsynaptic cell is depolarized beyond -40 mV.[2] This is because they lack surface AMPAR to pass current at hyperpolarized potentials, but do possess NMDARs that will pass current at more positive potentials (because of relief of magnesium block). Moreover, the EPSPs elicited with depolarized membrane potentials can be completely blocked by D-APV, a selective NMDAR blocker.[3]
Activation
Silent synapses are activated via the insertion of AMPARs into the postsynaptic membrane, a phenomenon commonly called "AMPA receptor trafficking."[4]
When glutamate binds to a strongly-depolarized postsynaptic cell (e.g., during Hebbian LTP), Ca2+ quickly enters and binds to calmodulin. Calmodulin activates calcium/calmodulin-dependent protein kinase II (CaMKII), which — among other things — acts on AMPAR-containing vesicles near the postsynaptic membrane. CaMKII phosphorylates these AMPARs, which serves as a signal to insert them into the postsynaptic membrane. Once AMPARs are inserted, the synapse is no longer silent; activated synapses no longer require simultaneous pre- and postsynaptic activity in order to elicit EPSPs.
Evidence suggests that dendrite arborization and synapse maturation 1 (Dasm1),an Ig superfamily member, is involved in the maturation of synapses, essentially "awakening" the silent synapses.
Controversy
The characterization of silent synapses is an ongoing field of research and there are many things about them that are not yet known. Some of what is currently accepted about the properties of silent synapses may still prove to be incorrect in whole or in part. Some controversies about silent synapses have however, been settled. For example, until recently, there were four competing hypotheses for the mechanisms of synapse silence:[5]
- The "whispering synapse" hypothesis:
- A synapse that releases glutamate more slowly than normal, thus activating only high affinity NMDA receptors, but not low affinity AMPA receptors
- The "low Pr" synapse hypothesis:
- A synapse that is not technically silent, but appears to be so, because it has such a low presynaptic probability of release that it rarely is activated.
- The "glutamate spillover" hypothesis:
- A synapse that does not release its own presynaptic glutamate, but in which the postsynapse detects low concentrations of glutamate "spilling over" from neighboring synapses. Only the high affinity NMDARs, but not the low affinity AMPARs can detect this low level of glutamate
- The "lack of AMPA receptor" hypothesis
- A synapse that lacks postsynaptic AMPA receptors
All four of these hypotheses had their adherents, but the first three were largely ruled out as a mechanism for synapse silence by work published before 2008.[6] However, recent experiments have clearly established that silent synapses can be observed at brainstem synapses bearing postsynaptic AMPA receptors.[7] This study favors the glutamate spillover hypothesis by showing that at silent synapses the glutamate concentration is reduced. At least, this study indicates that the popular hypothesis of the postsynaptic silent synapses does not apply in all systems.
See also
References
- ↑ Purves, Dale (2007). Neuroscience, Fourth Edition. Sinauer Associates. pp. 193–5.
- ↑ Liao, D.; Hessler, N. A.; Malinow, R. (1995). "Activation of postsynaptically silent synapses during pairing-induced LTP in CA1 region of hippocampal slice". Nature 375 (6530): 400–404. doi:10.1038/375400a0. PMID 7760933.
- ↑ Isaac, J. T.; Nicoll, R. A.; Malenka, R. C. (1995). "Evidence for silent synapses: Implications for the expression of LTP". Neuron 15 (2): 427–434. doi:10.1016/0896-6273(95)90046-2. PMID 7646894.
- ↑ Kerchner, G. A.; Nicoll, R. A. (2008). "Silent synapses and the emergence of a postsynaptic mechanism for LTP". Nature Reviews Neuroscience 9 (11): 813–825. doi:10.1038/nrn2501. PMC 2819160. PMID 18854855.
- ↑ Voronin, L. L.; Cherubini, E. (2004). "'Deaf, mute and whispering' silent synapses: Their role in synaptic plasticity". The Journal of Physiology 557 (Pt 1): 3–12. doi:10.1113/jphysiol.2003.058966. PMC 1665055. PMID 15034124.
- ↑ Montgomery, J. M.; Pavlidis, P.; Madison, D. V. (2001). "Pair recordings reveal all-silent synaptic connections and the postsynaptic expression of long-term potentiation". Neuron 29 (3): 691–701. doi:10.1016/S0896-6273(01)00244-6. PMID 11301028.
- ↑ Balland, B.; Lachamp, P.; Kessler, J. -P.; Tell, F. (2008). "Silent Synapses in Developing Rat Nucleus Tractus Solitarii Have AMPA Receptors". Journal of Neuroscience 28 (18): 4624–4634. doi:10.1523/JNEUROSCI.5355-07.2008. PMID 18448639.