Reversal potential
In a biological membrane, the reversal potential (also known as the Nernst potential) of an ion is the membrane potential at which there is no net (overall) flow of that particular ion from one side of the membrane to the other. In the case of post-synaptic neurons, the reversal potential is the membrane potential at which a given neurotransmitter causes no net current flow of ions through that neurotransmitter receptor's ion channel.[1]
In a single-ion system, reversal potential is synonymous with equilibrium potential; their numerical values are identical. The two terms refer to different aspects of the difference in membrane potential. Equilibrium refers to the fact that the net ion flux at a particular voltage is zero. That is, the outward and inward rates of ion movement are the same; the ion flux is in equilibrium. Reversal refers to the fact that a change of membrane potential on either side of the equilibrium potential reverses the overall direction of ion flux.[1]
The reversal potential is often called the "Nernst potential", as it can be calculated from the Nernst equation. Ion channels conduct most of the flow of simple ions in and out of cells. When a channel type that is selective to one species of ion dominates within the membrane of a cell (because other ion channels are closed, for example) then the voltage inside the cell will equilibrate (i.e. become equal) to the reversal potential for that ion (assuming the outside of the cell is at 0 volts). For example, the resting potential of most cells is close to the K+ (potassium ion) reversal potential. This is because at resting potential, potassium conductance dominates. During a typical action potential, the small resting ion conductance mediated by potassium channels is overwhelmed by the opening of a large number of Na+ (sodium ion) channels, which brings the membrane potential close to the reversal potential of sodium.
The relationship between the terms "reversal potential" and "equilibrium potential" only holds true for single-ion systems. In multi-ion systems, there are areas of the cell membrane where the summed currents of the multiple ions will equal zero. While this is a reversal potential in the sense that membrane potential reverses direction, it is not an equilibrium potential because not all (and in some cases, none) of the ions are in equilibrium and thus have net fluxes across the membrane. When a cell has significant permeabilities to more than one ion, the cell potential can be calculated from the Goldman-Hodgkin-Katz equation rather than the Nernst equation.
Mathematical models
The term driving force is related to equilibrium potential, and is likewise useful in understanding the current in biological membranes. Driving force refers to the difference between the actual membrane potential and an ion's equilibrium potential. It is defined by the following equation:
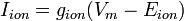 where
where 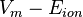 is the Driving Force.
is the Driving Force.
In words, this equation says that: the ionic current (Iion) is equal to that ion's conductance (gion) multiplied by the driving force, which is represented by the difference between the membrane potential and the ion's equilibrium potential (i.e. Vm-Eion). Note that the ionic current will be zero if the membrane is impermeable (gion = 0) to the ion in question, regardless of the size of the driving force.
A related equation (which is derived from the more general equation above) determines the magnitude of an end plate current (EPC), at a given membrane potential, in the neuromuscular junction:

where EPC is the end plate current, gACh is the ionic conductance activated by acetylcholine, Vm is the membrane potential, and Erev is the reversal potential. When the membrane potential is equal to the reversal potential, Vm-Erev is equal to 0 and there is no driving force on the ions involved.[1]
Use in research
When Vm is at the reversal potential (Vm-Erev is equal to 0), the identity of the ions that flow during an EPC can be deduced by comparing the reversal potential of the EPC to the equilibrium potential for various ions. For instance several excitatory ionotropic ligand-gated neurotransmitter receptors including glutamate receptors (AMPA, NMDA, and kainate), nicotinic acetylcholine (nACh), and serotonin (5-HT3) receptors are nonselective cation channels that pass Na+ and K+ in nearly equal proportions, giving an equilibrium potential close to 0 mV. The inhibitory ionotropic ligand-gated neurotransmitter receptors that carry Cl−, such as GABAA and glycine receptors, have equilibrium potentials close to the resting potential (approximately –70 mV) in neurons.[1]
This line of reasoning led to the development of experiments (by Akira Takeuchi and Noriko Takeuchi in 1960) that proved that acetylcholine-activated ion channels are approximately equally permeable to Na+ and K+ ions. The experiment was performed by lowering the external Na+ concentration, which lowers (more negative) the Na+ equilibrium potential and produces a negative shift in reversal potential. Conversely, increasing the external K+ concentration raises (more positive) the K+ equilibrium potential and produces a positive shift in reversal potential.[1]